
During the Fall of 2016, I was an undergraduate studying aquaculture at the University of Rhode Island. Like other aquaculture students, I was given an opportunity to work in one of the department labs as an assistant. It so happened, that I found myself employed by the Aquatic Pathology Laboratory under Dr. Marta Gomez Chiarri. My assignment was to assist a Ph.D. candidate in researching the interaction of a potential probiotic strain of Phaeobacter inhibins with the microalgae live feed Pavlova pinguis to combat the oyster pathogen Vibrio coralliilyticus.
Unfortunately, my first Ph.D. mentor left rather soon after I started, leaving me scrambling to learn what I could about microalgae culture, bacteria culture, and the processes of understanding the interaction of the two. Microalgae culture came to me quite naturally (autoclaves and lab incubators do not hurt), and the David Nelson Microbiology Laboratory upstairs did what they could to crash-course me through keeping various bacteria mutant strains alive (and separate). However, when it came to establishing a baseline understanding of microbial interaction, this paper was instrumental.
Simply put, this paper taught me to expect the unexpected when it came to microbial ecology. Much like relations between nations, microbes interact through the lens of economy, competition, and opportunity. Every relationship is contextualized by its environment. Unfavorable conditions can disrupt…and even destroy previously successful harmonies.
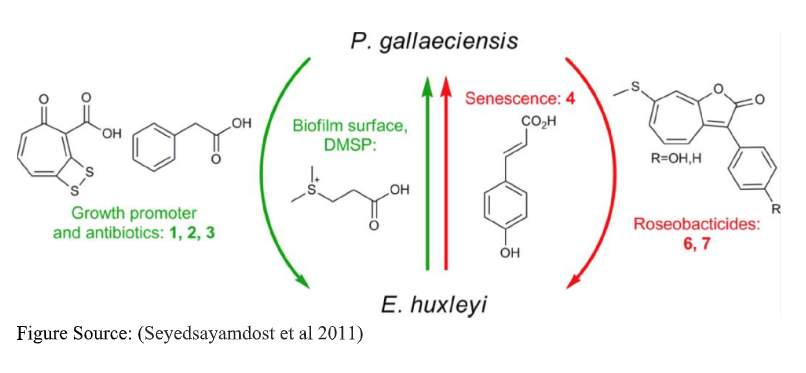
This paper by Mohammad R. Seyedsayamdost and team describes the changing nuances that come from any microbial relationship. This article focuses on the symbiosis of a pelagic diatom Emiliania huxleyi and Phaeobacter gallaeciensis. It is important to recognize that the natural life cycle of this diatom is one of ‘boom and bust’. A few sparse, atmospheric cells rapidly propagate once a supply of nutrients is upwelled by wind and wave action. During this ‘boom’, the diatom exhibits logarithmic growth which is, at this point, fueled by a microbial partner. Phaeobacter gallaeciensis provides Emiliania huxleyi with growth stimulants, in the form of auxins.

At the same time, the symbiont secretes localized antibiotics discouraging the growth of other microbes. This creates a domineering symbiotic ecology, one where mass amounts of Emiliania huxleyi bloom rapidly, hopped up on a constant flux of growth stimulants. The Phaeobacter receives two from this exchange. Likely localized sugars and oxygen secreted by the diatom host, but more importantly, space. Diatom tests (silicate shells) have immense amounts of complex surface area, perfect for the biofilm communities Phaeobacter likes to create. But this economy is precarious. For when the Emiliania huxleyi enter stationary growth phase, they begin to signal that resources have begun to become scarce and that growth should be minimized.
One symptom of an aging Emiliania huxleyi cell is p-coumaric acid, an algal lignin (fiber). This study demonstrated that once a threshold concentration of this ‘old man algae lignin smell’ is achieved, Phaeobacter turns from friend to foe. In the presence of p-coumaric acid, Phaeobacter gallaeciensis switches its metabolism from auxins over to the synthesis of algicides and rosebacticides. By these means, the once beneficial symbiont directly kills its host cells and likely disrupts/kills other associated beneficial microflora. The benevolent Dr. Jekyll has become a voracious Hyde. By such a transformation of metabolic character, Phaeobacter gallaeciensis becomes instrumental in controlling the immense diatom blooms itself helps create.
As I spend more time in the Reef Aquarium Industry, I see this paper as a means of rationalizing some of the sudden, confounding blights that sometimes befall well-established reef aquariums. Sometimes things are going so well for so long that disaster seems attracted to random targets. A specific coral’s flesh recedes. Zooxanthellae ‘decide’ to suddenly abandon another. A long healthy fish suddenly goes skinny. A sudden pest emerges even when nothing new has been added for months/years. All these things may indeed have a causation associated with shifts in microbial ecology…ultimately cascading from some environmental change (long or short term).
The issue with highly complex coral reef ecosystems is that such environmental changes usually do not collapse an established system. Complexity breeds robustness, yet if all is ‘well’ by the naked eye, long-term environmental changes can produce significant change over time. Changing environments can and will erode the stability of any system based on microbial ecology. Within every coral, fish, and invertebrate there is a careful dance between many bacteria and many host cells. The Jekyll and Hyde Chemistry of Phaeobacter gallaeciensis has taught us, with microbial ecology, to always expect the unexpected.
Literature Consulted
Seyedsayamdost, M. R., Case, R. J., Kolter, R., & Clardy, J. (2011). The Jekyll-and-Hyde chemistry of Phaeobacter gallaeciensis. Nature chemistry, 3(4), 331-335. Link to Full Article: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3376411/
About the Author
Taras Victorovich Pleskun is a professional aquaculturist originally from Mystic, Connecticut. Taras studied aquaculture at the University of Rhode Island and University of Florida. He has a MSc. in Aquaculture and specializes in the cultivation and application of microalgae. Taras has trained in aquaculture operations in the Philippines and Australia, experiences which changed how he appreciated the commercial farming of marine life. Taras is currently employed as Lead Phycologist at TopShelf Aquatics in Winter Park, Florida. His research interests include the application of microbes in reef aquariums, larviculture and sustainable feeds. Taras loves working and writing for the Reef Aquarium Industry. He truly feels that by learning to breed and appreciate all aquatic critters, humanity will be made smarter, stronger and kinder.
