
Rudie Kuiter is an Australian ichthyologist who has published extensively on Indo-Pacific reef fishes. Over the past half-century, he has described 56 new species, including a variety of wrasses, angelfishes, butterflyfishes, cardinalfishes, assessors, and, in particular, seahorses and pipefishes, though in recent years his attention has shifted to studying orchids and their pollinators.
His name will likely ring a bell for aquarists and divers thanks to several species that have been named in his honor—the damselfish Amblypomacentrus kuiteri, the leopard wrasse Macropharyngodon kuiteri, the dragonet Dactylopus kuiteri—as well as for the series of popular field guides that he has authored.
Some of these field guides are absolutely enormous in scope and feel more like taxonomic monographs, the most ambitious of which is his nearly 400-page magnum opus Labridae Fishes: Wrasses, published in 2010. In a review by Jake Adams, he rightly hailed this as the “Wrasse Bible”, and a 2nd edition would eventually be published by reefbuilders in 2015.
This work marks the first truly comprehensive treatment of the family (minus the parrotfishes) and incorporates a variety of taxonomic changes, including many new synonymies, resurrected taxa, reclassifications, and, most notably, a half-dozen new genera.
However, these changes would prove to be controversial within the scientific community. Kuiter arrived at his labrid classification by integrating morphological, biogeographical, ecological, and molecular data, the latter of which was based on several contemporaneous studies (Bellwood & Barber 2004, Westneat & Alfaro 2005, Cowman et al 2009) that had conclusively shown that certain large, cosmopolitan genera (e.g. Halichoeres, Coris) were in fact not natural groups, but rather were composed of species that had been lumped together based on superficial similarities of their morphology.
These troublesome genera were all members of the subfamily Julidinae, a group that has long confounded taxonomists, as can be seen by the plethora of discarded names lurking in the group’s convoluted taxonomic history.

To resolve this issue, Kuiter would resurrect several of these long-forgotten taxa (Iridio, Hemiulis, Platyglossus, Octocynodon, Hemitautoga, Hemicoris, Julis, and Pseudanampses), but he went even further by establishing new genera for nearly all of the undescribed clades (Biochoeres, Whitleychoeres, Corichoeres, Julichoeres, Paracoris, and Allocoris).
Owing to the similar morphologies of these fishes, the descriptive portion of this reclassification was necessarily terse, relying heavily on the existing molecular data to sort out their interrelationships. This isn’t to say that morphology was ignored, but the exhaustive osteological study that ichthyologists favor for these sorts of large-scale revisions was certainly nowhere to be found.
Not surprisingly, this irked some of the old guard in ichthyology, most notably the taxonomic demigod Jack Randall and his coauthor Paolo Parenti, whose 2010 labrid checklist dismissed Kuiter’s labors with a single sentence: “We do not accept this proliferation of labrid genera that lack morphological support.”, adding, “We concur that Halichoeres should be divided into several genera from a molecular standpoint. However, external morphological characters, including those of jaw and pharyngeal dentition, have not been found for such a division.”.
In the eyes of Parenti and Randall, a labrid genus that isn’t defined by its dentition is simply not worth bothering with, no matter what other supporting data might exist. Thus despite the overwhelming evidence that this family required extensive reclassification, Kuiter’s prescient work has languished for the past 14 years.
So neglected is the Wrasse Bible that you won’t even find mention of its new genera on the World Registry of Marine Species (WoRMS), as if these names simply didn’t exist, while the Catalog of Fishes defers to Parenti & Randall 2010, stating, “We tentatively do not accept these genera.”.
My dear reefbuilders, Rudie Kuiter’s wrasses are long overdue for a reappraisal, so let’s reappraise…
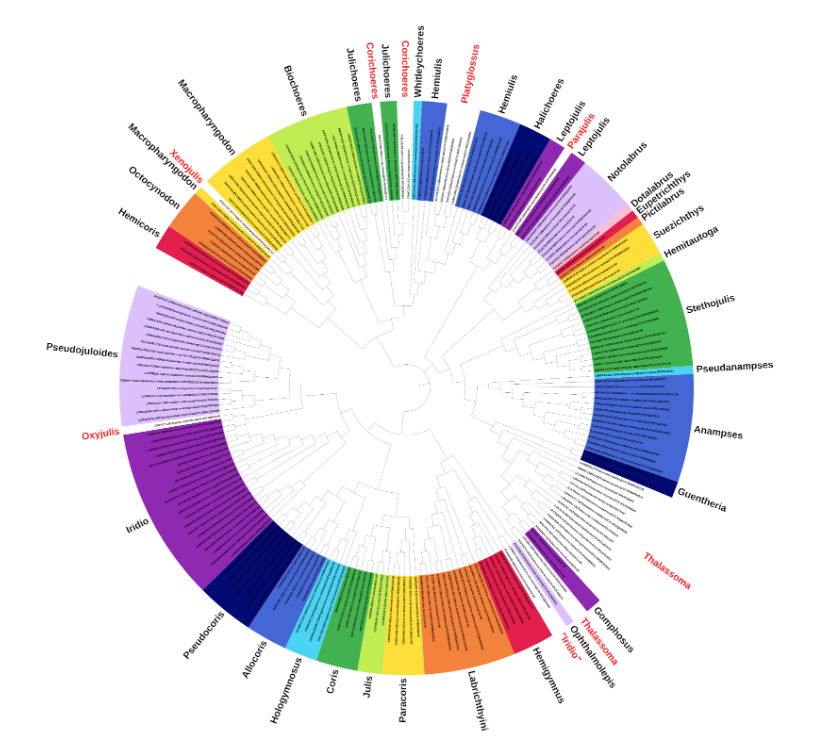
So what’s the verdict?
Nearly all of Kuiter’s resurrected genera appear to be supported (i.e. monophyletic): Hemicoris, Octocynodon, Hemiulis, Julis, and Pseudanampses. The one exception is Platyglossus, home to the melanurus and marginatus complexes (both of which are common in the aquarium trade); these appear to nestle together as a subclade within Hemiulis, all of which share a similar color pattern of horizontal green or brown stripes.
These are also vaguely similar to the 4 species in Octocynodon, which Kuiter suggested to be closely related, though this is argued against by the CO1 data, which instead places them on a basal clade with Hemicoris. The sole species in Pseudanampses, P. geographicus, sits as a closely related sister group to the more diverse Anampses, and is diagnosed by differences in its dentition and a greater number of lateral line scales, though whether these really need to be split is debatable.
When it comes to Kuiter’s newly described genera, we again find that all but one of these correspond to natural (i.e. monophyletic) groups: Biochoeres, Whitleycoeres, Julichoeres, Paracoris, and Allocoris.
The exception this time is Corichoeres, whose 2 recognized species were formerly included within Coris (owing to their high scale count), but which instead arise independently within the Julichoeres clade (diagnosed in part by a low scale count)—since both genera are described by Kuiter, these names have equal claim to the clade and would require revision to establish priority.
What’s intriguing about this group is their morphological and ecological diversity, including a mix of tropical and subtropical species, from both coral reefs and coastal habitats. These species do, however, share similarities in their juvenile color patterning, having an ocellus in the center of the dorsal fin and on the upper margin of the caudal peduncle.
The closely related Biochoeres and Hemiulis wrasses also feature a similar juvenile pattern but with additional posterior and anterior ocelli in the dorsal fin. On the other hand, the more distantly related Octocynodon lacks the caudal peduncle spot; such ontogenetic differences have largely been ignored when assessing the evolutionary relationships of this subfamily, but are likely to hold value. As they say, ontogeny recapitulates phylogeny.

Halichoeres hortulanus, which Kuiter removed to Hemitautoga, remains an enigma. The CO1 data fails to resolve its placement amongst the julidine wrasses, and finds inconsistent support for a relationship with its alleged congeners (sensu Kuiter), the Zigzag Wrasse H. scapularis and the Three-spot Wrasse H. trimaculatus.
These latter two species were once classified in their own genus, Guntheria, and it’s notable that their juveniles differ markedly from H. hortulanus, which instead has a peculiar 3-banded black & white patterning that’s unusual amongst the julidine wrasses, though vaguely similar to certain species of Macropharyngodon.
There are a few other foibles with Kuiter’s classification. The pair of species in Xenojulis are shown to have emerged from within Macropharyngodon, which isn’t surprising given the obvious similarities in their dentition and color patterning, the only major difference being the peculiar profile of the dorsal fin in Xenojulis.
Parajulis poecilopterus, a shoaling epibenthic species from the Northwest Pacific, falls within another genus of small, epibenthic fishes from the tropics, Leptojulis—all of these have a similar color patterning, with paired dark stripes, a dark pectoral blotch, and a dark y-shaped marking anterior to the dorsal fin… honestly, it’s hard to understand how these were ever regarded as separate genera.
There’s also the issue of Gomphosus and Thalassoma, home to such aquarium stalwarts as the Birdnose Wrasse and the Lunare Wrasse, respectively. In terms of morphology, these genera are essentially the same, save for the comically enlarged snout of Gomphosus, though this feature is only developed in mature specimens, with juveniles being nearly identical to some of the Thalassoma in terms of coloration.
This has been known about since the 2004 molecular study of Bernardi et al, but still nothing has been done to rectify the matter. The most sensible solution would be to sink Thalassoma into the older taxon, Gomphosus, but will anyone ever get around to doing so?
Lastly, there’s the matter of Iridio. The composition of this exclusively New World lineage has varied in several of the relevant molecular studies, with the most recent and comprehensive of these, Wainwright et al 2018, finding that all of the species fall into a single genus-level clade, with upwards of 5 distinct subclades that crossed over into the Eastern Pacific.
Included within this group are the formerly recognized genera Oxyjulis and Sagittalarva, plus a previously unresolved clade that included the Clown Wrasse Halichoeres maculipinna. A major sticking point in this group’s classification is the apparent discordance between the mitochondrial and nuclear data, with the former supporting polyphyly (i.e. multiple genera) and the latter supporting monophyly (i.e. a single genus, Iridio), but in either instance, we know with certainty that these are not Halichoeres, despite their ongoing inclusion within that genus.
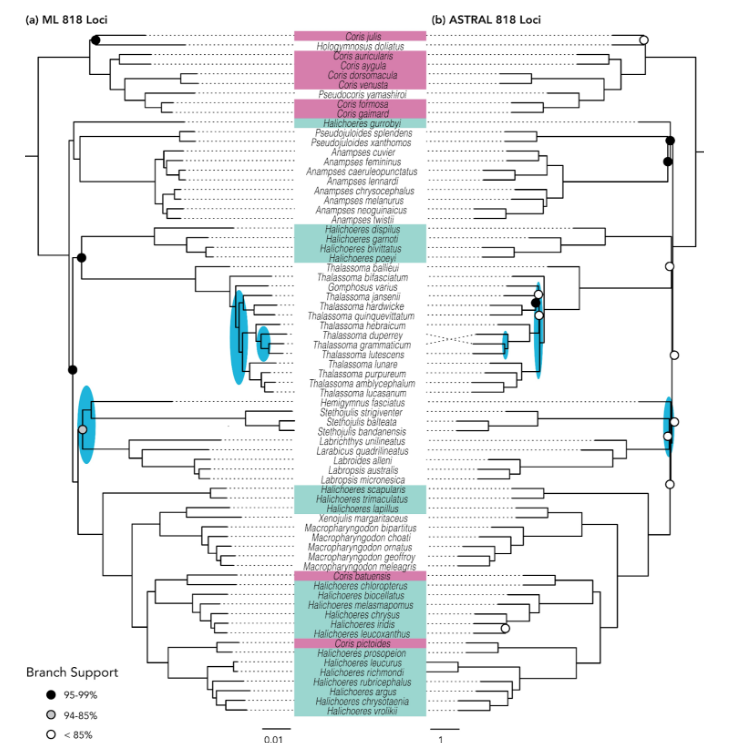
Recently, an even larger dataset appeared in the phylogenomic study of Hughes et al 2023, utilizing ~1100 exons and 300+ species of labrids. This provided a firmer footing for the deeper nodes within the labrid tree, resolving many relationships that had hitherto been uncertain, and amongst the julidines there was a general concordance with the layout of Kuiter’s reclassification, (though a few important lineages were absent from the analysis).
The question now is when will the inevitable happen. When will the julidine wrasses finally be split apart? The steady drumbeat of molecular discovery over the past two decades has all pointed in the same direction, and this has largely matched up with the revisions put forth in Kuiter’s Wrasse Bible. The only lingering uncertainty relates to how finely the julidines should be split.
What constitutes a genus-level lineage? With so little separating these groups in terms of morphology, should we instead define these by their inferred molecular age, and, if so, how old should a clade be to be considered taxonomically distinct: Oligocene… Miocene? For instance, the various Coris lineages that Kuiter split into a half-dozen distinct genera are similar in age to Thalassoma; certainly, the argument could be made that all of these Coris genera could be lumped together within a single large, heterogeneous genus.
Whatever its flaws and limitations, it’s become increasingly clear that Rudie Kuiter established a remarkably accurate labrid classification. Is it finally time to look past the reservations of Parenti & Randall and accept these necessary changes, or are we resigned to linger in labrid limbo indefinitely?
