

Let’s get deep in the weeds with fairy wrasse biogeography, specifically, the Cirrhilabrus cyanopleura group.
In 2022, Kai et al enlarged this group with the description of a pair of colorful new species, C. chaliasi and C. aquamarinus, both of which are commonly available in the aquarium trade and which had previously been confused with other similar species; this study also included the redescription of C. solorensis, a comparatively dull species that is rarely seen in captivity. This was followed up in 2024 with the addition of C. xanthozonus from West Papua, bringing membership in this group to 9 recognized species, all of which occur within the Andaman-West Pacific.
But there’s a conundrum with these fishes that might not be apparent until we plot them on a map. Look at Indonesia, absolutely overflowing with species-level diversity, most of which have small, restricted ranges (“microendemism”). When people point towards Indonesia as a center of marine biodiversity, this is what they’re referring to; certain groups seem to explode with a flourish of speciation, and the phenomenon is particularly prevalent within some of the species complexes in Cirrhilabrus. However, when we compare these fragmented distributions to C. cyanopleura, the northernmost member of this group, we encounter a sharp contrast. Over the years, opinions on this fish have varied, but it’s generally considered to be distributed from Japan to the Andaman Sea, south to Bali and east to Palau (records further east in Melanesia and Australia are erroneous). This is a vast range, particularly given the context of its closest relatives within the genus.
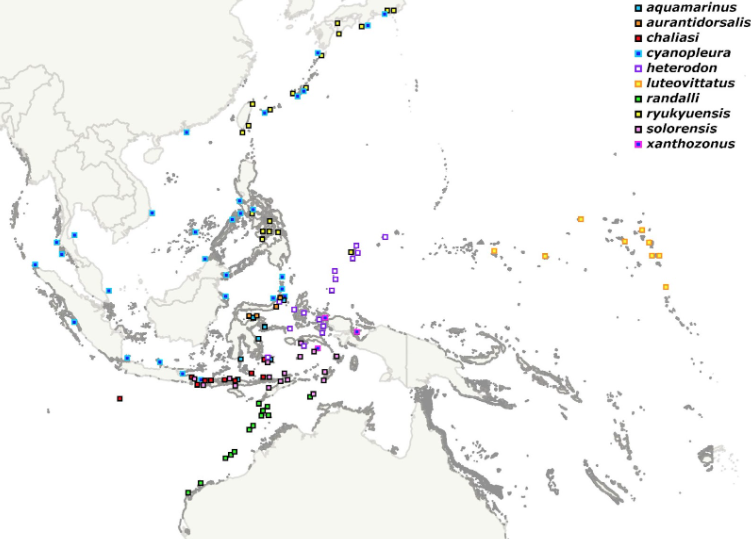
How do we explain this discrepancy? Why would cyanopleura succeed at maintaining its integrity as a species across such a vast expanse of the Coral Triangle when its closest cousins Balkanized themselves so thoroughly? Why did cyanopleura not also split itself along the various biogeographic boundaries that divide this region? Or have we gotten it all wrong? Is there evidence for speciation hiding in plain sight?
Now, the most obvious example of a cryptic species lurking within this group is the ryukyuensis phenotype, which is diagnosed by the presence of a yellow triangular blotch near the pectoral fins of males. On the surface, this seems like compelling evidence, but its validity has remained contentious. The species was described from Okinawa, where it is generally the dominant form, but the islands of the Ryukyu Arc and Japan are also home to the typical cyanopleura phenotype, albeit in lesser numbers, along with a wide spectrum of intermediate color morphs with varying amounts of yellow. In fact, it’s only in the central Visayan islands of the Philippines that we find ryukyuensis existing as a more or less fully allopatric population (i.e. behaving as a proper species should), though as it ranges further north it does eventually encounter and interbreed with cyanopleura in places like Romblon. So is ryukyuensis a species?
To answer that, we first need to more accurately define what we mean by cyanopleura. The species is the second oldest in the genus, taxonomically speaking, described from Jakarta by the eminent Dutch ichthyologist Pieter Bleeker in 1851. Topotypical males feature a blue “hood” that symmetrically covers the anterior third of the body, but the yellow pectoral blotch seen in ryukyuensis is always lacking. We can follow this same form south to Bali (where it is very rarely encountered, replaced instead by its red-headed relative, C. chaliasi), and north into the Andaman Sea, the Gulf of Thailand, and further east into the South China Sea, where the powerful Kuroshio Current guides them upwards towards Japan. Viewed from this biogeographic lens, we can begin to envision a Pleistocene world in which a species occurring on the western shores of Sundaland (cyanopleura) is periodically isolated from its sister species to the east (ryukyuensis), only for these to find themselves reintroduced when the glaciers melted and sea levels rose, repeatedly allowing for a renewal of gene flow between them.
But what of those populations found further south, at Sulawesi and West Papua and Palau? When and how did cyanopleura successfully cross the major biogeographic boundary known as Wallace’s Line that separates the Philippines from Indonesia, and, more importantly, how was gene flow preserved once this happened? After all, on a geologic time scale, species are unlikely to maintain themselves in the absence of gene flow.
Rudie Kuiter presents an alternative hypothesis in his Labridae Fishes: Wrasses from 2010, arguing that C. cyanopleura is in fact endemic to the Eastern Indian Ocean and the Java Sea, replaced elsewhere in the Pacific by a broadly defined C. ryukyuensis, present from Japan to Palau and encompassing a variety of phenotypes that may or may not possess a colored blotch near the pectoral fins. This is not as unreasonable as it might sound given how many other labrids split along these same biogeographic lines, but his revision is problematic for a couple reasons. 1) Kuiter’s heterogenous mix of phenotypes within ryukyuensis is difficult to reconcile when compared to the various microendemic species found just to the south. 2) There is an older name available in the literature that would have priority over ryukyuensis for such a widespread species: Cirrhilabrus heterodon.

In addition to describing C. cyanopleura, Pieter Bleeker also named the closely related C. solorensis from Solor, Indonesia in 1853 and C. heterodon from Ambon, Indonesia in 1871. But in retrospect, Bleeker had only a cursory understanding of these fishes; his descriptions were based on single specimens and relied on traits that we would now consider to be uninformative (pelvic fin length & dentition), though as he had the advantage of being one of the first ichthyologists to study this region’s fauna, most of his species descriptions, no matter how poorly diagnosed they may be, have stood the test of time.
The end result was that Bleeker’s Cirrhilabrus taxa have been (and continue to be) poorly understood. L. F. de Beaufort, in his The Fishes of the Indo-Australian Archipelago from 1940, made the first attempt at reclassifying the group, recognizing solorensis and cyanopleura as valid species, though he erred in following Bleeker’s usage of pelvic fin length (a trait that develops in males of this group) to delimit these species, thus misidentifying some of his specimens. And so when he wrote, “C. heterodon, the type of which I have examined in the Leiden Museum, is certainly the same as C. cyanopleura”, it’s not entirely certain which species he actually compared it against, as he was simply acknowledging that the subadult phenotype is similar amongst these fishes. This synonymy has been blindly followed ever since, by the likes of Randall, Allen & Erdmann, and Kai et al 2022, but to date there has never been any meaningful analysis of the various cyanopleura populations, either morphological or molecular. But we can attempt to address that right now…

There are in fact a few subtle differences that become apparent when comparing topotypical specimens of C. cyanopleura versus C. heterodon. We’ll focus on the fully developed male phenotype of these fishes, as younger specimens within this group all look more or less the same. Firstly, the blue “hood” of cyanopleura is demarcated posteriorly by a straight vertical line, whereas the hood in heterodon has an oblique posterior margin, with the underlying coloration of the body encroaching forwards towards the head, revealing darkly margined scales that are otherwise hidden in cyanopleura. C. heterodon is also a bit more colorful on its sides, sharing a similarly orange dorsum with cyanopleura, but with a distinct teal patch anterior to this, with additional regions of lavender and aquamarine pigment often present below. The color of the hood also differs slightly near the pectoral fins, having a greenish or bluish tone in cyanopleura (alluded to in its name), while in heterodon this varies from a greyish-brown to a dull purple. That being said, exceptions abound amongst these fishes, particularly within the biogeographical melting pot around Sulawesi, and so it’s not uncommon to encounter specimens with aberrations that render them indefinable. Hybridization is expected to be rampant in areas where multiple species occur in sympatry, such as in the Lembeh Strait.
Special mention needs to be made of the population in Palau, which exhibits a confounding mix of phenotypes alongside a few unusual traits. In a minority of specimens there is a broadly developed and evenly colored blue hood similar to that of cyanopleura, while in others the hood is distinctly oblique and has a contrasting teal patch below the dorsal fin. Unique to this population is the development of an opaque patch of pigment posterodorsally, which conceals the underlying dark scale margins and iridescent markings (compare this to specimens from Indonesia, which have the dark scale margins visible through the orange dorsum); such opaque pigments also occur in the dorsum of C. aurantidorsalis and the sides of C. luteovittatus, C. randalli, and C. xanthozonus, illustrating the importance such markings have in the speciation of this group. The base of the dorsal fin also has a similar opaque color through most of its length, whereas cyanopleura and heterodon typically have only the anteriormost interspinous membranes translucently yellowed, with the remainder containing a dark (likely fluorescent) pigment. The purpose of these opaque pigments is to conceal the underlying fluorescence that members of this group possess; likewise, the blue hood of cyanopleura and heterodon serves a similar function, whereas cranial fluorescence is prevalent in the unhooded species.
Palau sits at the western edge of the Central Pacific, with its marine fauna composed of species from both Indonesia and Micronesia; however, recent evidence has shown that there is also potentially some small amount of connectivity with the southern islands of the Philippines. In videos shared by Bruce Carlson and photos from Keoki Stender, specimens of the yellow-blotched ryukyuensis phenotype are documented here, along with the unique orange-backed Palauan phenotype. This amalgamation of otherwise disjunct species may explain the heterogenous appearance of this population, formed from a mix of cf heterodon, ryukyuensis, and their various hybrids.
All this is to say that, yeah, C. heterodon is very likely a valid species endemic to a narrow portion of Indonesia, from Ambon to Wakitobi, north to Lembeh and Raja Ampat, with similar specimens from Palau potentially representing another undescribed sister species. It gets trickier when we try to define the western boundary of heterodon, though the Lembeh Strait, home to the closely related C. aurantidorsalis and C. aquamarinus, is probably about as far as it gets, as the hydrology in this region generally suppresses migration further westward. On the other side of Sulawesi’s Minahasa Peninsula, at Bunaken, the population is mostly of the cyanopleura phenotype, as are specimens along the eastern edge of the Celebes Sea at Bangka Island and the Sangihe Islands. Heading further west to Borneo, cyanopleura is again prevalent, but Kuiter notes that specimens at Mabul have a “more pinkish” pectoral blotch, though this population remains a bit too poorly documented to draw any firm conclusions.
So then where does C. cyanopleura actually occur? Is it endemic to the Andaman and Java Seas, as Kuiter would have it? Are the populations from the Gulf of Thailand and South China Sea conspecific with it? What about specimens from Palawan and Luzon? And how about those that occur together with the ryukyuensis phenotype throughout the Ryukyu Arc? And what about those isolated populations in Northeastern Borneo and the Celebes Sea? Are these all one big messy cyanopleura that speciated in isolation and subsequently spread to fill these disparate regions, or are there multiple cryptic species involved with indiscernible male phenotypes? When we consider the endemism prevalent within this genus (and in particular within this group), it’s tempting to consider the latter as the more likely hypothesis, but heaps of molecular data will be needed before we have a fuller understanding of their evolutionary history.
And what of ryukyuensis? Is it a valid species, or is it perhaps a variable phenotype of the northerly cf cyanopleura population whose proportion changes geographically? Why does it occur in isolation within the central islands of the Philippines but in conjunction with cyanopleura throughout its type locality in the Ryukyu Arc? Does this imply that it speciated in the Philippines and later spread northwards or vice versa? There are, in fact, a few other examples of reef fishes with this same peculiar Okinawan-Visayan endemism, most notably the One-spot Foxface Siganus unimaculatus and to some extent the Magenta Dottyback Pictichromis porphyrea, but the phenomenon is certainly uncommon. This latter example is particularly interesting, as it also includes populations in Palau, Ambon, and east to Micronesia (those from Fiji are likely an undescribed species), thus mirroring in some respects the cyanopleura group. Is it possible that both groups emerged in Micronesia and later spread to the West Pacific? This would deviate from the presumed evolutionary history of most marine life in this region, but it’s not implausible.
Deciphering the species limits of the cyanopleura group will be an enormous undertaking, but the lessons learned from solving this puzzle have implications for resolving some of the other outstanding taxonomic mysteries in the Indo-Pacific. In the aquarium trade, C. cyanopleura is heavily exported from Java and the Philippines, but specimens of C. heterodon are also likely to find themselves collected in smaller numbers from Sulawesi. For the fairy wrasse connoisseur, it’s a rarely seen species worth keeping an eye out for.

Main image credit [Cirrhilabrus heterodon, from Misool, West Papua. Credit: Sabine Templeton]

{kind=link}