
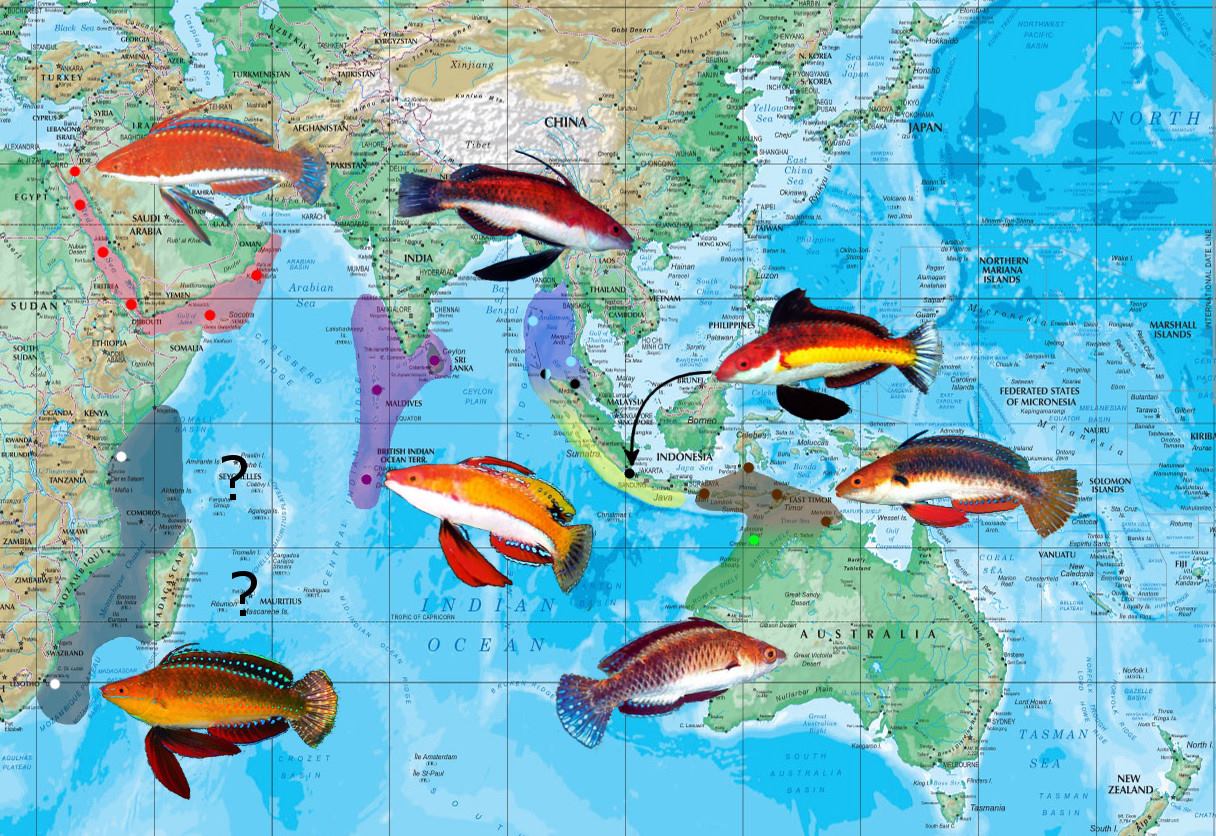
The rubriventralis group features some of the most beloved and charming species of fairy wrasses, ornately adorned with painterly brilliance and morphological exuberance. Unlike some of the characteristically larger members of Cirrhilabrus, the rubriventralis group members remain small, rarely exceeding three inches in standard length. The group is entirely Indian Ocean in distribution, with two main populations being distributed on each side of the Ninety East Ridge along the 90th meridian. Cirrhilabrus rubriventralis and its geographically distinct phenotypes occupy the Western Indian Ocean, while the East Indies play host to four highly similar species distributed along the Sunda-Banda Archipelago.

Springer and Randall described the first species from this group in 1974, from specimens obtained from the Red Sea. Only seven other Cirrhilabrus were known prior to this, making it the 8th official member to this then rapidly growing genus. The new fairy wrasse was noted for having remarkably large pelvic fins, in which they were bulbous, trailing, and entirely red. It was subsequently named Cirrhilabrus rubriventralis, after this unique morphological feature.
Allen described Cirrhilabrus morrisoni and C. joanallenae as the 2nd and 3rd members of this group nearly three decades later, in 1999 and 2000 respectively. He obtained specimens of both species from the Timor Sea and Northwestern Sumatra respectively, and noted similarities in the pendulous pelvic fins. Nine years later in 2009, Randall and Tanaka described Cirrhilabrus naokoae based on males obtained from the aquarium trade. In 2012, Allen and Erdmann described Cirrhilabrus humanni from Alor, making it the 5th and newest scientifically recognized species in this group.
Like most other groups, however, we see various isolated populations in familiar biogeographic regions of endemism, suggesting the presence of possible cryptic species. This is seen particularly in C. rubriventralis, where the African and Maldivian populations are already showing clear differences in phenotype.
It’s interesting to note that while the species are easily diagnosed via color and morphology, they are virtually indistinguishable based on preliminary molecular analysis. Commonly used genetic markers such as CO1 sequences often show negligible variances between the members, suggesting that the rubriventralis group is at the incipient stages of their evolutionary divergence, where genetic change is greatly outpaced by a phenotypic one. As such, the phylogeny of this group remains largely unresolved, with the proposed tree being left as a polytomy.

With such a level of genetic homogeneity between the speciated taxa, it would seem more than likely that hybridization should run rampant in this group. Ironically, hybridization does not occur not because of potential genetic incompatibility, but rather from the lack of geographical contact. The rubriventralis group members are largely allopatric, with only one pair confirmed to overlap narrowly on the northernmost tip of Sumatra, in the province of Banda Aceh. And as expected, hybridization is thought to occur here.
Members of this group are some of the easiest Cirrhilabrus to diagnose. As with keeping with this lineage, males of all species possess very long pelvic fins, but these are modified into comically pendulous, bulb shaped extensions. This trait is highly unique, and is shared with no other species outside of this group. Another characteristic feature of the rubriventralis group is the presence of a pennant on the anterior dorsal fin. These are formed by the extrapolation of the first and second dorsal spines, and in males, are elongated to considerable filamentous extensions. Females lack this dorsal fin embellishment, possessing instead a stubby bulb. This bulb only extends into the trademark pennant during transition into the terminal male phase, and is therefore a secondary sexual characteristic brought about under the influence of testosterone. This applies to the pelvic fin development as well. Only three other species in the filamentosus clade of the genus Cirrhilabrus possess dorsal filaments, but they differ by having those present anteromedially of their dorsal fins.
A metallic spot is present on the base of each interspinous membrane on the dorsal fin, and is arranged linearly so as to form a dotted stripe. This is another unique and important feature in diagnosing the group, and is shared with members of the Cirrhilabrus condei clade, supporting our assertion that these two diverse lineages are actually geminate sister clades divided by the Sunda shelf. The soft dorsal fin also possesses a series of medially placed spots, somewhat reminiscent of those seen in the temminckii group. Although these are never well developed enough to form the distinct medial macular stripe seen in the temminckii group or pylei clade, its presence hints of a shared relationship and warranted placement in this lineage. All members lack a visible dorsal stripe, but possess lateral stripes that ranges from being highly vestigial, to clearly defined. Facial stripes are mostly absent, but are occasionally displayed as vestigial or unobtrusive manifestations.

Their caudal fins are weakly double emarginated, and display an incredible luster unmatched by any other Cirrhilabrus species outside of this group. At peak nuptial activity, the caudal fin takes on a metallic foil-like resemblance, is extremely iridescent and is capable of reflecting large amounts of light. This trait is assumed to be homologous with members of the rubrimarginatus, temminckii and filamentosus groups, and supportive of their common ancestry within the genus.
The males exploit this incredible caudal scintilla during their nuptial displays, and they do so by very quickly paddling their tails from side to side in a rudder like fashion. This novel nuptial behavior is restricted to the rubriventralis group members only and is a unique evolutionary behavior most probably brought upon by sexual selection.

The females are largely similar between species, and are fairly useless in diagnostics. They are uniformly orange to red, fading to white ventrally, possessing a single black spot on the caudal peduncle. Females from the rubriventralis group possess fine spots and striae, presented in a series of horizontal stripes along the dorsum and lateral body. Depending on the species and developmental stage, these range from being muted to highly iridescent. The pelvic fins and dorsal filaments are undeveloped and therefore uncharacteristic at this phase.
All the rubriventralis group members are very susceptible to aquarium induced darkening, where the intense red dorsum takes on a sooty black shade. The intensity of aquarium lights may have a part to play in this, which is curious considering these fish occur in fairly shallow, well-lit reefs.
Cirrhilabrus rubriventralis (Red Sea [Type locality])

In this attractive species, the ground color is uniformly bright red, variably tinted in orange. The ventral half is white, and the two colors are delimited neatly in place of an absent lateral stripe. Occasionally, this intangible stripe is represented as a row of metallic spots. This lateral stripe, while inconspicuous in the resting phase, becomes very prominently displayed in metallic scintilla during nuptial flashing. The body is not uncommonly lightly marred in a fuliginous grey. Cirrhilabrus rubriventralis lacks any obvious facial stripes, but these may appear as vestigial manifestations in certain individuals.
The dorsal fin is red but heavily obfuscated in black, especially toward the posterior soft dorsal region. Each membranous space between the rays are presented with a single metallic spot, placed at the base of the dorsal fin in a horizontal row so as to form a dotted stripe. A second row of spots are placed medially along the posterior soft dorsal fin, but are not developed enough to form the characteristic macular banding seen in the temminckii group. The first and second dorsal fin spines are greatly elongated to form a filamentous pennant, in which the males retract and erect at will for courtship.


The scarlet pelvic fins are pendulous and exaggerated, trailing past the anal fin in fully developed males. This is undoubtedly the most characteristic feature for this species, and is shared with all members of its group. The anal fin is red and unmarked, except for a row of intermembranous spots at the base. The caudal fin is weakly double emarginated, finely spotted and uniformly steely blue.
No photos of nuptial males from the Red Sea are currently known, but in all likelihood, it shares a similar display pattern as those from adjacent regions such as the Maldives. The dorsal fin is folded against the body while the dorsal pennant is raised. The red body lightens to a bright white, and the lateral stripe becomes very apparent in metallic blue. The caudal fin takes on an incredible metallic luster, and at certain angles becomes capable of reflecting light.
This phenotype is known from the Red Sea and the adjoining Gulf of Aden up to Oman. Curiously, despite the region being heavily dived and explored, very few photographs of this species are available in literature. This is translated in the aquarium trade as well, where it is practically absent. Cirrhilabrus rubriventralis in the aquarium trade are almost exclusively collected from Sri Lanka and the Maldives.
The history of the Red Sea is written through multiple glacial events with corresponding rising and falling of sea levels. This has resulted in the separation of the once homogenous population of Indian Ocean species, catalyzing speciation via selection and isolation. Therefore, despite this phenotype being the type representation for the species, it is quite likely that it stemmed from a larger population outside of the Red Sea, possibly from the neighboring Maldives. The Maldivian phenotype is therefore hereby treated as a geographically and phenotypically distinct cryptic “species”.
Cirrhilabrus cf. rubriventralis (Maldives, Sri Lanka & the Chagos Archipelago)

This phenotype is essentially identical to the preceding, but can be separated based on a few differences in color. The ground coloration is more variably plastic, and ranges from intense red to a light yellow-orange. The posterior soft dorsal fin is almost always yellow, unlike the fuliginous red of the type representations. All other aspects of this phenotype remain the same, including the vestigial nature of the lateral and facial stripes, and the reddening of the pelvic fins.




This phenotype is distributed across the Sri Lankan-Maldives-Chagos ecoregion. It is strangely absent from the Seychelles and Mauritius, although it should be expected to occur there. If it does, its biogeography should also most likely result in some form of phenotypic difference. This is the commonest member of the rubriventralis group to occur in the aquarium trade. Its exuberant color, comical finnage and relatively inexpensive nature make this one of the most popular Cirrhilabrus amongst amateur hobbyists. It is, however, fairly aggressive and particularly susceptible to darkening under intense lighting.
Cirrhilabrus cf. rubriventralis (Africa)

The African population of this species is currently known only from Kenya and KwaZulu-natal, but it more than likely occurs throughout the East African coast between both regions. It harbors considerable differences in coloration from the type species, as well as the Maldivian and Sri Lankan phenotypes. Terminal males are a rich fulvous orange dorsally, fading to a lighter tawny amber toward the ventral region. The facial stripes are more prominently present, being represented as a metallic stripe above and below the eye.
The lower facial stripe continues to extend towards the caudal peduncle as a series of linearly placed spots. This is essentially a modified lateral stripe, and in this phenotype, is mostly present and very distinct. A series of shorter stripes comprised of the same linearly placed metallic spotting are present above the lateral stripe as well. The dorsal fin is mahogany, and spotted in the same fashion as with the other rubriventralis complex members. Like C. rubriventralis, a pennant is present on the anterior dorsal fin.
The anal and pelvic fins are blood red, and in the former, is decorated with a row of metallic spots at the base. Unlike the steely blue in C. rubriventralis, the caudal fin of this phenotype is distinctively cuprous, reflecting accents of pink and gold at various angles just like elemental copper would.






This phenotype assumes the standard posture during nuptial display. The entire dorsal fin save for the pennant is contracted and pressed against the body, and the metallic spots glow in a scintillating brilliance. The coppery sheen on the caudal fin intensifies as well, reflecting various shades of pink and gold. This phenotype, although not rare from East African exports, cannot be considered common. Like all other members of the group, it has a particular proclivity for aquarium induced darkening.
The distinct difference in phenotype might suggest a more advanced stage of speciation as compared to the neighboring population of C. cf rubriventralis in the Maldivian archipelago. However, despite the obvious physical differences, mitochondrial CO1 sequences show minimal genetic variation from C. rubriventralis. This is in keeping with the group, where members typically show a greater discordance in color amongst designated species than genetics. This raises a point regarding the consistency in how taxonomists treat the various species in this group. A similar scenario is presented in Cirrhilabrus joanallenae and Cirrhilabrus naokoae. Both species share identical CO1 sequences, but the latter has been awarded species level based solely on differences in color and morphology. Going by this principal, should Cirrhilabrus cf. rubriventralis from the East African coast not follow suit?
Cirrhilabrus joanallenae

Cirrhilabrus joanallenae is very similar to Cirrhilabrus rubriventralis in terms of coloration. The two can be separated easily based on the following. 1) In C. joanallenae, the pelvic fins are always jet-black versus red in C. rubriventralis. 2) The interface between the red dorsum and white ventral body is variably traced in yellow for C. joanallenae, but this feature is never present in C. rubriventralis. 3) In C. joanallenae, the caudal peduncle region is heavily dusted in black, whilst in C. rubriventralis this region is usually clean. The most reliable diagnostic feature still lies in the differences in pelvic fin coloration. All other phenotypic aspects of this fish remains similar to that of C. rubriventralis.



Despite its small geographical distribution, Cirrhilabrus joanallenae shows remarkable variation in the extent of its yellow lateral stripe. This feature ranges from completely absent, to copious, and does not appear to correlate with any particular geographical location. The variability of this stripe foreshadows Cirrhilabrus naokoae, to which C. joanallenae is perhaps most closely related to.

In nuptial flashing, C. joanallenae adopts the typical posture of clamped fins save for the dorsal pennant. The dorsoanterior portion of the body just beneath the pennant lights up in white, while the rest of the dorsum lightens to a bright yellow-gold. In typical fashion of this group, the caudal fin scintillates in an electrifying steely blue.
Cirrhilabrus joanallenae is distributed toward the northern tip of Sumatra, from Weh Island in Banda Aceh, to Phuket in Thailand and the Andaman Sea. It mixes narrowly with Cirrhilabrus naokoae in Banda Aceh, where they likely hybridize. The species was named in honor of Gerry Allen’s mother, Joan Allen, and is Latinized in the epithet “joanallenae”.
Cirrhilabrus naokoae

This is without a doubt the most interesting and beautiful member of the rubriventralis group. Cirrhilabrus naokoae is red on the dorsal portion of the head, covering the upper lip, eye and upper portion of the operculum flap. Below this demarcation is a stark white, and the interface is partitioned by the presence of very vestigial facial stripes, one above and below the eye. The red continues across the entire dorsum to a demarcation from above the pectoral fin base to midbase of the caudal fin, where it is sparsely suffused in a smoky black. A broad yellow belt runs equatorially just below the red dorsum, starting at the pectoral fin base and ending at the ventral half of the caudal fin base. The pectoral fin base is red within this yellow belt. The ventral body is stark white, giving the fish a tri-colored appearance.
The white ventral belly and inferior portion of the head are on different planes, with the facial delimitation being almost continuous but not connected with the yellow equatorial belt on the medial body. As such, a white panhandle is seen protruding at the preoperculum region. This feature is important, and serves to differentiate it from Cirrhilabrus joanallenae and the hybrids that the two form. This will be discussed in greater detail at the end of the article.

The dorsal fin morphology for this species is unlike any other in the genus or its group. The first four dorsal spines are elongated, tapering down gradually in height before continuing with the rest of the posterior dorsal fin. This results in an upright lobular pennant that is very unlike the wispy, filamented examples seen in the other rubriventralis group members. The pelvic fins are pendulous and large, and are featureless black. The caudal fin is weakly double emarginated, and has the usual steely blue hue.

In nuptial display, the fins are clamped down in the usual fashion, except for the pennant on the anterior dorsal fin. Because C. naokoae’s dorsal fin pennant is lobular and triangular, the raising of it results in a shark-like appearance as it cruises around its habitat. The face and posterior section of the dorsum lightens to a downy white, and the caudal fin takes on the usual remarkable scintillating brilliance.
The embedded video above shows C. naokoae in nuptial colors. Take note of the caudal reflectance and the incredibly unique swishing motion it incorporates into its display. Cirrhilabrus naokoae was described from three specimens obtained via the aquarium trade. For a long time, its exact geographical distribution was not known, but narrowed down to plausible locations along Sumatra. Because fish collection in the Indonesian archipelago is rife with shoddy and unregulated rigmarole, pin pointing a species to the exact collection source is often fraught with immense difficulty. Today, this species is confirmed to exist from Jakarta, Medan and north to Banda Aceh. It mixes very narrowly with Cirrhilabrus joanallenae in the lattermost region, where the two species hybridize.

Mitochrondrial CO1 sequences for both species have yielded essentially no differences, and so C. naokoae was described solely based on color, morphology and meristics. Their biogeography suggests that both species are sisters, especially taking into account the variability of the yellow stripe in C. joanallenae. In C. naokoae, this feature is essentially just a slightly more prominent and permanent feature. Cirrhilabrus naokoae was named in honor of Dr. Hiroyuki Tanaka’s wife, Naoko. Tanaka was the co-author for the species’ description.
Cirrhilabrus naokoae is a fairly recent addition to the genus and the aquarium scene, debuting in 2008. Since then, it has quickly become a staple, and the fish is now quite commonly obtained from Indonesian imports. The species is pugnacious and has the typical proclivity for darkening in captivity.
Cirrhilabrus humanni

Cirrhilabrus humanni is the newest member of the rubriventralis group, being officially described in 2012. The species is variably colored along the sides, having a dingy orange base color overlaid by extensive patches of red and brown mottling. This shading can range from nascent to highly prominent, at times completely obscuring the underlying orange. The head is reddish, and delimited from the whitish ventral region just below the eye. Terminal males completely lack any form of lateral or facial stripes, although these are present in the usual lineated spot patterns in females and transitioning males.
The pelvic fins are large and pendulous, as with all other members of this group, but unlike C. naokoae, C. joanallenae and C. morissoni, are completely red and unmarked. This is more in fitting with Cirrhilabrus rubriventralis and its geographically distinct phenotypes in the Western Indian Ocean, versus the former three with black pelvic fins in the Eastern Indian Ocean. This makes C. humanni the only species in the East Indies in possession of red pelvic fins.


The dorsal fin is uniformly red, with the first two anterior spines elongated to form a pennant. The dorsal pennant in Cirrhilabrus humanni is characteristic, elongating only very slightly, before terminating into a spade. This has coined the species with the moniker “Stubby-fin Fairy Wrasse”. However, the feature is also variable, and depending on the individual, may present with a barely noticeable clubbed appendage. A row of spots are present between the membranous spaces along the base of the dorsal fin, and a second row of spots run parallel along the medial portion just above.
The anal fin is likewise red, delimited in white at the base. The interface is lined with a row of metallic spots in the same fashion as the dorsal fin. The caudal fin is of the usual steely blue, but with underlying tones of olive.

During nuptial displays, the fuliginous mottling dispels to reveal a rich fulvous body. No visible lightening or metallic iridescence is observed to develop. The fins are clamped down in the usual fashion, save the prominent erection of the anterior dorsal pennant. Here, the spade shaped elongation that gives the species its namesake becomes very apparent. The tail scintillates in a sterling silvery-blue, and reflects light as the male swishes it from side to side at incredible speeds like a rudder. Again, this novel behavior is something restricted to members of this group, and is not seen anywhere else.



Cirrhilabrus humanni is restricted to the Lesser Sunda Islands, with patchy and scattered distribution records. The type specimen was captured from Alor, Indonesia, and was thought to occur only there at the time of description. The species is not uncommon in East Timor, where it occurs in typical rubble pans at depths of 20-25m. It has also been found northwest in Batuata, south of Sulawesi. A single aquarium specimen was traced back to Northern Bali; however, whether or not it was actually collected there or simply exported from there remains unclear. Bali exists as a dubious record for now.

In August of 2015, an unknown Cirrhilabrus was captured in Darwin, Northern Australia. The photograph above was sent to me for identification, and was subsequently forwarded to Gerry Allen and Fenton Walsh. It was later identified as Cirrhilabrus humanni, with Darwin representing a new range extension for the species. This is the southern limit for the species across the Timor Sea, and in Northwestern Australia, Cirrhilabrus humanni is replaced by the very similar Cirrhilabrus morrisoni.
Cirrhilabrus humanni is very rare in the trade, with few, sporadic records of specimens entering from Indonesia. It has the same behavior in captivity as with the other rubriventralis group members, including the penchant for darkening under intense lightning. This species was named in honor of Paul Humann, who first sighted this fish with Ned and Anna DeLoach in Alor, Indonesia.
Cirrhilabrus morrisoni

In this species, the ground coloration is peachy orange fading to white ventrally. As with Cirrhilabrus humanni, the body is heavily obfuscated in a fuliginous black, but this feature is more clearly defined in C. morrisoni. The sooty obfuscation comes from the darkening of each scale, the extent of which determines the phenotypic variability amongst individuals. This also results in an argyle like pattern, especially on the lighter anterior body. It lacks any noticeable lateral or facial stripes.
The dorsal fin is black, and lined on the base with the usual row of metallic spots placed in between each membrane. The first and second spines are elongated slightly to form a pennant, but in this species, is short and fairly straight. The pennant is of similar length and form to C. humanni, but lacks the spade like clubbing at the terminus. This diagnostic is often unreliable in separating the two, as C. humanni sometimes lack its characteristic clubbed pennant amongst individuals. The most reliable key for separation lies in the coloration of the pelvic fins and the biogeographical distribution of the two.

The pelvic fins are large, pendulous, and featureless black. The anal fin is black, delimited in white at the base, and the interface is lined with a row of spots similar to those of the dorsal fin. The caudal fin is of the usual steely blue, decorated in a series of spots and striae.
Because of this species’ isolated range and inaccessible nature, not many photographs of it exist. Likewise, not much is known about the species, including its nuptial coloration. It is expected to follow the typical pattern seen in this group, with fins clamped down and pennant raised, as well as lightening of the body and metallic iridescence developing on the caudal fin.


Cirrhilabrus morrisoni is restricted to the northwestern Australian region, specifically in Hibernia Reef. It shares this unique biogeography with three other Cirrhilabrus species, namely C. randalli, C. cf. temminckii and C. cf. exquisitus. These four are essentially geographically isolated representations of their respective groups; with C. randalli and C. morrisoni achieving species level allocation. It is both surprising, and unsurprising, that the temminckii and exquisitus representatives here are still treated as synonyms of the existing populace. This serves as another example of the inconsistent treatment of taxa across multiple groups in this ubiquitous genus.
Another species occurring here is Conniella apterygia. This species was covered briefly in the lanceolatus group article, where the morphological lack of pelvic fins was hypothesized to stem from a genetic bottleneck due to its highly isolated and restricted range. It remains curious, however, that C. morrisoni and the other endemic Cirrhilabrus in this region are able to maintain their form despite the same environmental conditions presented to Conniella. Could the lack of pelvic fins in the latter therefore be attributed to something else? These are questions that are unanswered for now.
C. morrisoni is found in rubble pans replete with Halimeda. Because of its isolated and inaccessible range, this species is not available to aquarists, and will likely not relinquish its title of “book fish” anytime soon. C. morrisoni was named in honor of Hugh Morrison, owner of the Perth Diving Academy in Australia.
Hybrids
While the bulk of Cirrhilabrus hybridization occur in the cyanopleura group, we’ve seen various examples being documented in the lubbocki, lunatus, scottorum, exquisitus as well as temminckii groups. The rubriventralis group presents one documented case of hybridization between Cirrhilabrus naokoae and Cirrhilabrus joanallenae in the northern region of Sumatra, specifically in Banda Aceh and Medan.

Despite the similarities in genetics between C. joanallenae and C. naokoae, the two are characteristically distinct enough to separate via phenotype. The contrasting differences result in hybrids with an even expression of traits from both parents, making identification fairly straightforward. In the example above, an intermediate dorsal fin pennant between both parental species can be observed. In C. joanallenae, the pennant is formed by the elongation of the first two dorsal spines, and is extremely thin, wispy, and hair like. The pennant in C. naokoae is markedly different, being sail like, involving the first four dorsal spines instead. The hybrid shows intermediary traits of both, with a significantly shorter and thicker pennant than those seen in C. joanallenae and C. naokoae.

While both C. joanallenae and C. naokoae possess a lateral yellow belt at the red and white body interface, the stripe is always weak and highly variable in the former, but strong, thick, and very clearly defined in the latter. Hybridization of the two species often results in an intermediary phenotype where the yellow belt is suffused and indistinct. Such is seen in the example above. Another character distinguishing C. naokoae from C. joanallenae is the presence of a white preoperculum panhandle. In the hybrid above, this feature is vestigial, but visible enough to suggest a blood tie to C. naokoae.
Because both species are genetically very similar, standard genetic sequencing will fail to yield any usable information. Mitochondrial CO1 gene sequences of C. joanallenae and C. naokoae are identical, and so are those between C. rubriventralis and its African counterpart. This genetic homogeneity is likely to occur in all members of the group, where separation of species have traditionally been carried out based on the sole premise of morphology, biogeography and color. This incipient genetic divergence is therefore unlikely to be seen via usage of standard genetic markers, requiring perhaps nuclear DNA, or the comparison of longer, more comprehensive selection of genetic material.
Ironically, this homogeneity is not reflected in morphology, and despite the early stages of speciation, the various group members are easily and very readily separated based on appearance. To the west of the 90th meridian in the Western Indian Ocean, we see Cirrhilabrus rubriventralis in the Red Sea, as well as two geographically isolated phenotypes in the Maldivian-Chagos Archipelago and the East African coast. Both variants show distinct phenotypic differences from the type species, especially the one from Africa, where elevation to species level may be warranted. The Maldivian population is presumably closest to the Indonesian taxa, since Africa and the Red Sea are highly distinct ecoregions in most examples of reef fish biogeography.
To the East of the 90th meridian in the East Indies, four species are currently scientifically recognized — all, with the exception of C. humanni, are in possession of black pelvic fins. Only C. joanallenae and C. naokoae are sympatric, where their hybridization is discussed above.
In summary, the rubriventralis group is fascinating for the many reasons presented above. The discordance in genetic versus phenotypic divergence in a group at the cusp of evolution could be attributed to the interplay between biogeographic isolation and sexual selection. This is further supported by the novel behavior used by males in nuptial display, where their caudal fin is swished repeatedly back and forth like a rudder. The rubriventralis group sets the stage for the discussion of the filamentosus group – the final group in the long-finned clade, and the penultimate discussion prior to Cirrhilabrus laboutei, as well as the end of this taxing taxonomic review.
