
The filamentosus group presents the reader with a collection of familiar, aquarium-friendly species, many of which are exuberantly colored and morphologically quirky. Despite some superficial differences in colors and fin shapes, the overall morphology strongly implies that this is the Pacific counterpart to the Indian Ocean rubriventralis group. Given the highly diverse and endemic nature of both groups, as well as the considerable differences between the Indian and Pacific Ocean taxa, it seems these fishes have an unusual propensity and rapidity when it comes to speciating.

Cirrhilabrus filamentosus was first known in 1976 from specimens collected in Indonesia. The species was described just two years after C. rubriventralis, making it the second species of Cirrhilabrus known to possess a dorsal filament. C. filamentosus, however, had an anatomically thicker and more pronounced filament formed from the extension of dorsal spines 10 and 11, versus the thin and wispy extensions of spines 1 and 2 in C. rubriventralis. Four years later, in 1980, Randall and Carpenter described Cirrhilabrus rubripinnis from the Philippines. Cirrhilabrus condei was later known from New Guinea in 1996, followed by Cirrhilabrus tonozukai and C. walshi in 1999 and 2001 respective. Despite the numerous descriptions throughout the years, no real phylogenetic relationships have been drawn between the members.
In 2015, Allen, Erdmann and Dailami split Cirrhilabrus condei into two species based on differences in morphometrics and coloration. The new species was named Cirrhilabrus marinda, after Drs. Marcus Wanma and Drs. Inda Arfan (Marcus and Inda = marinda), the Bupati and Vice Bupati of Raja Ampat. However, in review of the publication, we feel that the split was not strongly supported, and in this article, we treat C. marinda synonymously with C. condei. This will be discussed in greater detail later on.

Members of this group possess at the base of their dorsal fins, a row of metallic spots placed linearly between each spine and ray, a feature shared with the rubriventralis group. Their pelvic fins are long, often trailing past the anal fin origin, and the caudal fins are lustrous, possessing scintillating scales when viewed in a sidelight. This feature is very prominently displayed during courtship. The throat region of all species possess blue scaling of varying extent, at times covering the entire gular region in metallic blue.
The handful of species can be broken into a pair of informal subgroups that correspond to their fin shape and biogeography. Those found in Indonesia and the Philippines – referred to here as the filamentosus clade – possess a strong filament placed anteromedially along the dorsal fin. These are formed from the elongation of spines 10 and 11, and are capable of trailing past the caudal fin in the largest of males. It is of importance to note that C. rubripinnis lacks this feature, but occasionally, specimens are seen in possession of a tiny vestigial filament, hinting at the closeness and justified placement of the species in this clade. The members lack any distinctive dorsal, facial or medial stripes, but the lower belly is often delimited in white or cream in place of the usual medial stripe. Unlike the rubriventralis group and condei clade, the anal fins are largely unmarked, sans a few metallic spots placed randomly toward the posterior edge. Their pelvic fins are long and trailing, are distinctively leaf shaped and intermediate in width between those of the rubriventralis group and the condei clade.

The aforementioned condei clade is comprised of a pair of species, which range throughout Melanesia, as well as a questionably described third species. They lack any dorsal fin embellishments. Instead, the anterior spinous half arches significantly higher than the posterior soft dorsal fin, lending a sailfin like appearance to the males. This is especially evident during nuptial displays, when the anterior half is raised in full prominence, while the posterior half is folded down against the body. Their caudal fins are, in keeping with this lineage, very reflective, but in members of the condei clade the central rays of the caudal fins are particularly lustrous, forming a distinct strip strongly delimited from the rest of the fin This feature is unique within the genus, but it is best observed in person, as photographs fail to fully capture the ephemeral nature of this characteristic. The dorsal lateral line stripe is absent here, but there is the indication of a medial stripe that begins beneath the eye and extends posteriorly. This is sometimes poorly seen in resting males, but becomes very evident in nuptial display.


Females of this group are characteristically unique, but are fairly useless in separating down to species level. All are brick red, possessing on the caudal peduncle a single black spot, and a white spot of similar size at the tip of the snout. The dorsum and medial body are lined with fine striae of bluish white. This is very similar to those seen in the rubriventralis group females, but, in the latter group, the striae are more prominent, metallic, and confined to the dorsum. Nonetheless, the similarities yet again suggest a shared relationship between both groups.
The filamentosus clade

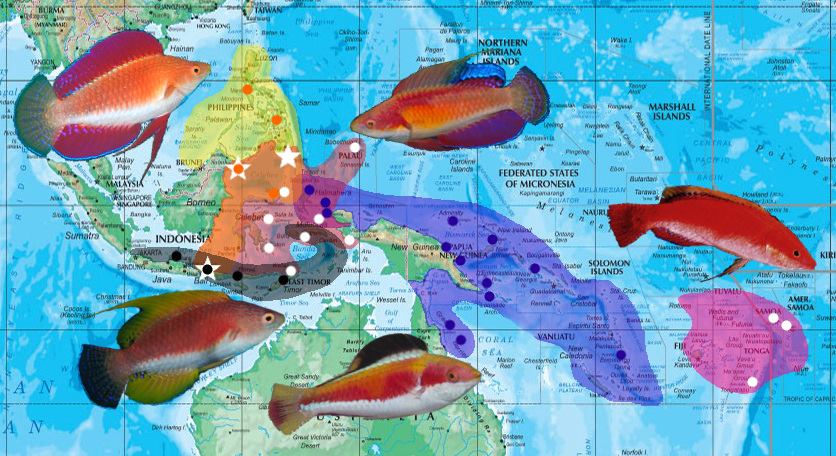
The three species in this group have neighboring biogeographies restricted to Indonesia and the Philippines. This limited range is unique in the genus, as there is no northward extention into regions like the Ryukyu Islands of Japan or the waters north of Borneo. This is in stark contrast to the widespread distribution of the rubriventralis group in the Red Sea and Indian Ocean, though why these two sister groups should differ to such an extent is a bit of a mystery.
The contiguous ranges seen within the filamentosus clade, as well as with the condei clade, creates expanses of overlap which allow for hybridization (and possibly even hybrid speciation, in the case of Cirrhilabrus tonozukai). Such hybrids have caused a great deal of taxonomic confusion, but they are more easily understood in light of their occurrence in these interspecies contact zones. There is still a great deal of uncertainty regarding the precise pattern of their speciation, so we will leave the matter unresolved until proper genetic study is available to elucidate the matter.
Cirrhilabrus filamentosus

Cirrhilabrus filamentosus is an undeniable poster species of the Indonesian Archopelago. In this iconic Cirrhilabrus, the males are uniformly brick red dorsally and medially, fading to a whitish ventral region. The delimitation is marked without a medial stripe, but is clean and clearly defined. The head and dorsum is occasionally striped in numerous hair fine striae, but these are often seen only in females and subterminal males. The feature diminishes in prominence in terminal phase males. The species lacks any dorsal and facial stripes. The gular, or chin region, is scaled in metallic blue, the extent of which varies amongst individuals.
The dorsal fin is uniformly mustard and arched significantly higher at the anterior region before trailing off in a robust and thick filament formed from the fusion of dorsal spines 10 and 11. This filament is often capable of trailing well beyond the caudal fin in the largest males, earning the species its common name of “Whip Fin Fairy Wrasse”. Each membrane space between the dorsal fin rays is marked at the base in a single metallic spot. This trait is seen in all members of the filamentosus and rubriventralis groups and is a homologous characteristic that ties the two closely. The posterior soft dorsal fin is usually unmarked but occasionally sports random occurrences of indistinct spots and streaks.
The anal fin is marked in the same manner and color as the dorsal fin. The pelvic fins are long, mustard yellow and broadly leaf shaped, although they are never as pendulous as those seen in the rubriventralis groups. The caudal fin is round, steely blue and riddled with a byzantine scrawling. As in keeping with all the species in this lineage, the caudal fins exhibit a reflective scintillation, reaching its peak reflectance during nuptial displays.


In nuptial display, Cirrhilabrus filamentosus lightens to a fulvous orange over the entire head, chest and dorsum above the medial delimitation. The white ventral belly darkens to a sooty obsidian, save for a small anteriomedial blotch in white. This blotch is much like what we see in the nuptial patterning of C. balteatus and C. temminckii, hinting at the shared ancestry of all these diverse wrasses. Cirrhilabrus filamentosus appears to lack any development in scintillating stripes or body patches during its display; however, its caudal fin now takes on a reflective, cellophane-like quality in rich cobalt. The pelvic, dorsal and anal fins are held against the body, and it is in this position that the whip-like filamentous dorsal extension can be truly appreciated.


Cirrhilabrus filamentosus is distributed throughout the Banda Archipelago from Bali to East Timor, where it is abundant. The species occurs in large groups of mainly females, over areas of extensive rubble patches with the occasional Euphyllia or xeniid coral. It is almost always found in the company of Paracheilinus species, namely P. cf. filamentosus and P. flavianalis. Males often spar with each other by swimming parallel against one another with fins fully extended and gular flap puffed out. The pelvic fins are held out and can be flicked independently. Hybrids presumably involving this species have been documented in Bali, but this requires further investigation that will be discussed in greater detail toward the end of this article.
Cirrhilabrus filamentosus is widely available in the aquarium hobby where it retails at very reasonable prices. Because of its colorful nature, affordability and quirky fin morphology, the species is always attractive to the novice aquarist. However, they will soon learn that C. filamentosus is a species of belligerent nature, capable of heinous crimes of insidious intent towards other fairy wrasses.
Cirrhilabrus rubripinnis

Cirrhilabrus rubripinnis is another handsome species of timeless beauty. In this classic fairy wrasse, the body is marked the same as with the preceding species, being tomato red instead of the usual fulvous orange. The ventral region is of a lighter shade, sometimes white, and is smoothly delimited without the presence of a medial stripe. The usual blue metallic scaling is present on the gular region. No dorsal or facial stripes are present in this species.


The dorsal fin is deep maraschino red, featureless, and raised significantly at the anterior spinous portion. The base is decorated with the usual spots present in each membrane space, but this feature is often obfuscated. The dorsal fin contour follows that of Cirrhilabrus filamentosus, but, instead of terminating in a filament, it makes a sharp dip before continuing with the posterior dorsal fin. This gives the fin a unique sail-like morphology. In very rare instances, C. rubripinnis is capable of sporting a filament in the usual position, but this is always severely reduced and presumably a result of atavistic expression.


The soft dorsal fin is inconsistently marked with a constellation of metallic blue spots. This varies from absent, to weakly present, and in some specimens is capable of forming a continuous band along the medial portion of the dorsal fin. The anal fin is marked the same but is edged in metallic blue on the outer margin.
The deep red pelvic fins are long, leaf-shaped and broad. The caudal fin is yellow, and is inscribed with the usual squiggling. Despite the nondescript caudal fin coloration, the scintillating qualities are still present, but only seen during the peak of nuptial display.


As with all Cirrhilabrus species, males have varying states of excitation levels during nuptial display, at which point the color changes accordingly. In full nuptial display, the head and chest region lighten to a rich fulvous orange. The blue scales beneath the gular region scintillate in a deep cobalt, extending to the base of the pelvic fins. The body lightens to light orange, with the addition of a white saddle developing just behind the pectoral fin. This saddle stretches from the dorsal fin to the pelvic fins, and is capable of bleeding in suffusion along the rest of the body. However, the head always maintains its distinctive orange during this stage.
The fins take on a magenta slate, and the caudal fin turns opaque crimson with a steely sheen of fuchsia varying in reflectivity according to angles of incident light. This is where the homologous trait really shows in prominence, closely associating it members of this lineage.


Cirrhilabrus rubripinnis is distributed in the Philippines down to Northern Sulawesi. Unidentified, presumably hybrid, phenotypes have been documented in the Southern Philippines, Sulawesi, and, curiously, Bali. It’s not exactly clear what species are involved in the formation of these unusual hybrids, but, should C. rubripinnis be involved, it would make Bali the southernmost point of its distribution. This would likely occur as exceedingly rare waifs along the Makassar Strait, which links Bali and the Philippines via a strong southerly current.
Cirrhilabrus rubripinnis is common and affordable in the aquarium trade, where its gaudy coloration and flamboyant finnage are greatly attractive to the novice aquarist. The species can be belligerent and feisty, but usually not to the extent of the preceding species.
Cirrhilabrus tonozukai

Cirrhilabrus tonozukai is quite possibly the most interesting member of this clade. In this species, the ground coloration is tangerine orange, fading to a lighter shade ventrally in a smooth suffusion. The head is loosely coifed in red, delimited to just below the eye and extending to the pectoral fin base and the anterior part of the dorsum. The usual blue scaling is present in the gular region, reaching down to the pelvic fin base. The species lacks any dorsal and medial stripes, but possesses one stripe just below the eye. This is usually obfuscated, becoming prominent only during nuptial display.


The dorsal fin is distinctively taller at the anterior spinous half, much like those in C. filamentosus and C. rubripinnis. A single filament emerges from the ends of spines 10 and 11, but is much shorter than those of C. filamentosus. It rarely, if ever, reaches past the caudal fin. The dorsal fin is translucent orange, except for the posterior soft region. Here, it is marked with an oblong, horizontally placed medial spot in red. The spot is outlined in black, and enveloped in a lattice network of ramifying blue squiggles. As with all members of this group, the base of the dorsal fin is lined in a series of metallic spots, each placed within the membranous spaces of the fin.
The pelvic fins are red. The anal fin is magenta and largely unmarked, sans the posterior portion where, occasionally, individuals sport haphazard manifestations of blue spots. The caudal fin is translucent orangey-red to yellow and possess the usual scrawling seen in this clade. It only displays the scintilla during nuptial displays.

In nuptial display, C. tonozukai adopts a uniformly fulvous body. The region immediately behind the pectoral fin base lightens to a downy white, but this never forms a saddle like that seen in C. rubripinnis. At peak excitation, this white patch can extend toward the caudal peduncle. The red coif is reduced to a streak behind the eye to the operculum, and another tracing the dorsum. This darkens to a deep crimson, becoming very stark and noticeable. The blue scaling at the gular region shines with iridescence.
The dorsal fin reduces in opacity and takes on a hyaline appearance, while the posterior squiggling and the basal spots scintillate in metallic blue. In keeping with this group, the caudal fin darkens to an opaque magenta, complete with a reflective shimmer.


Cirrhilabrus tonozukai is distributed primarily around Sulawesi, east to Raja Ampats, Halmahera and Palau, although populations there are likely to be vagrant and rare. A 2013 survey of Timor Leste revealed the presence of this species, making it the southernmost record of its distribution. Here, it is photographed cavorting with C. filamentosus and C. humanni. A handful of unnamed, possibly hybrid phenotypes that strongly suggest the genetic involvement of C. tonozukai have been documented in Bali and southern Philippines, suggesting that this species is capable of extending out of its usual range as rare larval waifs.

It is also curious to note that C. tonozukai occupies a parapatric distribution, with C. rubripinnis, C. filamentosus and C. condei bordering its northern, southern and eastern limits respectively. If one were to remove C. tonozukai completely, the biogeography of the remaining three species would portray a near perfect allopatric jigsaw. It might not be entirely implausible that the distributions of these species started converging slowly over the last half-million years, resulting in some contact where C. tonozukai currently occupies. Could C. tonozukai therefore arise from the speciation of some ancestral hybridization of its sister taxa? It would not be entirely contentious, seeing as the species sports some plasticity in its phenotype, especially that of its variable dorsal filament. It also appears to bear some intermediate phenotypes of its adjacent congeners, but, without extensive molecular studies, this hypothesis will serve to remain only as such.
C. tonozukai is relatively uncommon in the aquarium trade, making sporadic appearances from Sulawesi imports, usually with the likes of C. aurantidorsalis. The species is affordable and slightly belligerent.
The condei clade

Unlike the filamentosus clade, the condei clade members lack any filamentous embellishments on their dorsal fins. They do, however, still possess the prominent heightening of the anterior dorsal fin, before sharply continuing with the posterior soft dorsal fin. Like the members of this group, and those of the closely related rubriventralis group, all members possess the usual row of spots present at the dorsal fin base.
The condei clade is unique for possessing an additional chatoyant strip on the central caudal fin rays. This is extra lustrous in comparison to the rest of the fin, often taking on a magenta or reddish sheen in a sidelight. This unique apomorphy is not shared with any other Cirrhilabrus groups, and serves to distinguish the condei clade members with extreme ease.
The clade traditionally houses two members – C. condei and C. walshi. In early 2015, C. condei was split based on a cryptic species diagnosed with apparent differences in fin coloration and morphometrics. The split yielded Cirrhilabrus marinda, a largely sympatric species that occupies different niche habitats. In this case, C. marinda prefers offshore oceanic reefs, while C. condei prefers more inshore habitats. In examining the two phenotypes closely, we feel the diagnostic characters listed by Allen et al 2015 are unreliable and ignore many examples of intermediate phenotypes. For this reason, we’ll be treating marinda as a junior synonym of condei.
These species range throughout Melanesia, from the Raja Ampats to Fiji and Samoa, and south into the Great Barrier Reef. Specimens might reasonably be expected to turn up in New Caledonia as well, though none have yet been recorded there.
Cirrhilabrus condei

In Cirrhilabrus condei, the ground color is uniformly ruby-colored, delimited sharply to a white ventral region with a single, continuous medial stripe. The stripe starts from below the eye, ending at the caudal peduncle. The dorsal fin is raised anteriorly and is deep crimson. The usual row of metallic spots is present at the base of the fin. The margin of the anterior dorsal fin is variable in height and coloration, from almost fully black to only marginally so. The posterior soft dorsal fin is always rimmed on the outer edge in yellow, and this is delimited from the crimson ground color with a row of medially placed spots.
The anal fin is similarly marked, but without any contrasting colors on the outer edge. The caudal fin is lustrous yellow and marked extensively with metallic spots and streaks. The central rays are steely magenta, forming a distinct longitudinal strip. The pelvic fins are long, trailing past the anal fin, but are never as broad as those in the filamentosus clade.


In early 2015, Cirrhilabrus condei was split based on alleged differences in the dorsal fin structure and coloration, as well as perceived differences in habitat preferences. It is noted to possess the following:
1) The anterior dorsal fin is significantly taller and more “sail-like” in Cirrhilabrus marinda versus that of Cirrhilabrus condei, which supposedly has a more uniform profile. This difference is reflected in the mean maximum dorsal spine height, which is 15.2% of the SL (range 13.0-16.7, n=11) for C. condei and 19.5% of the SL (16.9-22.7, n=13) for C. marinda (Allen et al, 2015).
2) The black region on the dorsal fin of C. marinda is significantly darker, with the black region engulfing nearly the entire anterior spinous portion, tapering well into the posterior soft dorsal fin. Conversely, in Cirrhilabrus condei, this black region is restricted to the outer margin, never reaching to the base of the dorsal fin and is about equal widths throughout the length.
3) The nuptial coloration differs between C. marinda and C. condei. In the former, the anterior spinous portion of the dorsal fin is held erect and in a sail-like manner. This is apparently not seen in the latter. In C. condei, the posterior segment of the medial stripe is more prominently displayed, as well as the presence of extra white ornamentations along the eye, pectoral and posterior body not seen in C. marinda. The presence of a yellow patch also serves to differentiate the latter from the former.
4) C. marinda is also reported to attain a smaller maximum size than that of C. condei.

However, in analysis of various specimens from photographs taken in the wild and in captivity, most of these supposedly diagnostic traits are somewhat capricious, occurring and overlapping in both “species”. For example, the specimen depicted in Fig. 1 shows a significantly taller anterior dorsal fin profile, which is suggestive of Cirrhilabrus marinda; however, the restriction of the black coloration to the periphery of the dorsal fin is more indicative of C. condei. In Fig. 4, the male specimen sports the extensively black dorsal fin characteristic of C. marinda, but it lacks the taller anterior spinous portion. Instead, the fin exhibits a rather even profile, which is supposedly typical of C. condei.
These characteristics appear to run the gamut between individuals, seemingly counter indicative of the diagnostics between the two “species”.

The differences in nuptial coloration are also not consistent and reliable in support of C. marinda’s status as a valid species. In Fig. 3, the differences in nuptial patterns for C. marinda and C. condei were noted in the following. 1) The latter possesses extensive ornamentations of white behind the eye, pectoral and medial regions of the body. 2) The former holds the anterior portion of the dorsal fin erect while the latter does not. 3) The latter has, on the posterior dorsum, an extensive yellow patch which is otherwise absent in the former.
Fig. 5 above shows a specimen in nuptial display yet again in possession of traits seen from both “species”. The erection of the anterior dorsal fin coupled with the extensive black coloration is suggestive of C. marinda, while the white body markings and yellow posterior dorsum is indicative of C. condei.


It’s quite clear from these photos that the diagnostic phenotypic allocations for C. condei and C. marinda are not exclusive to the two and therefore are not reliable keys for separation. The fluidity in phenotype could be attributed to regular intraspecies variation, and differences in nuptial coloration could be influenced by numerous factors, some of which include state of excitation, as well as maturity of displaying males. It isn’t uncommon for certain species to possess varying levels of pigmentation on a specific anatomical feature. Cirrhilabrus cf. bathyphilus, for example, displays the same variability in its dorsal fin pattern.

It is also noted that Cirrhilabrus marinda frequents oceanic reefs situated further offshore, with higer levels of current and wave action, as opposed to Cirrhilabrus condei, which frequents sheltered, inshore reefs (though specimens of C. condei collected in the Coral Sea have come from the same type of oceanic habitats alleged to be favored by C. marinda, futher blurring their distinction). This difference in niche habitats could account for the supposedly marked difference in sizes of the two. C. marinda is said to attain a much smaller maximum length in parts of its range, which may be due to environmental factors of its habitat affecting abundance of food. Perhaps an inshore, sheltered reef would be more beneficial in aiding growth and development, as compared to a population living in turbulent, oceanic waters. If this is true, then the difference in size is unlikely to be caused by speciation, but more in line with that of an environmental cause.
The wide sympatry between the two species is also somewhat curious, with no other examples of sister Cirrhilabrus displaying such a unique biogeography. The genetic analysis of their mitochondrial CO1 gene sequences showed no differences in the two as well. There is indeed a precedent for coral reef fishes being recognized as distinct taxa despite having identical CO1 sequenes (e.g. Pseudojuloides edwardi vs. P. severnsi), but it has never been used to justify splitting entirely overlapping populations before. As should be demonstrably clear from this series, the dominant driver of speciation for Cirrhilabrus is biogeographic isolation.
Ultimately, we have chosen to conclude that this marinda phenotype is likely nothing more than normal variation within condei, given that: 1) the morphological characters separating condei/marinda are ambiguous and prone to sampling bias. 2) The known genetic sequences are statistically identical. 3) The fish is poorly known in the wild and Coral Sea populations offer equivocal support for there being any real habitat distinction. 4) The sympatric biogeography is entirely anomalous within the genus. While we have chosen a conservative approach here, more genetic study is warranted to resolve this issue.




Both phenotypes are sympatric, but the black finned “marinda” phenotypes have only been found in Halmahera, West Papua, the Bismark Archipelago and Vanuatu. Cirrhilabrus condei occurs in the same regions, and in addition, is also found in the Solomon Islands, Papua New Guinea, Coral Sea and the Great Barrier Reef. Since both phenotypes are likely to be synonymous, it is quite likely that both forms are found in all regions within this distribution range. If ever there were cryptic species to be found within C. condei, Australia and Vanuatu might be potential places to start looking, seeing as those regions are notorious for housing endemics.
Cirrhilabrus condei is moderately common in the trade, obtainable from Australian and Vanuatuan imports. It is fairly inexpensive, relative to some of its congeners coming from the same region. C. condei makes for an excellent aquarium fish, maintaining its colors very well with a proclivity for frequent nuptial displays.
Cirrhilabrus walshi


Cirrhilabrus walshi is essentially identical in appearance to the preceding, with one major difference – the dorsal fin. In C. walshi, the dorsal fin is entirely yellow, lacking any black pigment near, or on the peripheral edge. A medial band comprised of small, laterally placed spots run along the length of the dorsal fin. In C. condei, this is restricted only to the posterior soft region beneath the yellow margin.
Cirrhilabrus walshi is a lesser-known species, with no nuptial coloration ever being documented. The species represents an allopatric sister of C. condei, and is known from American Samoa, where it inhabits reefs at depths ranging from 30-60m. The species is named in honor of Fenton Walsh, who first collected the type specimens from Tauma Banks. He reports seeing this species in the company of Cirrhilabrus scottorum.

C. walshi is an archetypical “book fish” – a species out of reach to aquarists, known only from literature. This species should be expected to occur in nearby Fiji and Tonga, and aquarium collectors should be on the lookout for this species in moderately deep waters around rubble pans.
Hybrids

The hybridization of this group centers around three species in the filamentosus clade, namely C. rubripinnis, C. tonozukai and C. filamentosus. The biogeography of these species puts C. tonozukai in close contact with the other members at the edge of its range. We therefore expect to see hybridization occurring here.
The specimen above displays intermediate phenotypes of C. tonozukai and C. rubripinnis. The blotch on the posterior dorsal fin, for example, is a trait clearly passed on from the former. It’s interesting to note that in this hybrid, the gular and pelvic region is extensively scaled in blue, a trait that is only very sparingly present in both parental phenotypes. The locality of this specimen from Sulawesi suggests C. tonozukai and C. rubrpinnis to be the parental precursors, seeing as C. filamentosus does not range this far north into Sulawesi. C. rubripinnis is rare here, occuring at the edge of its distribution.

The photo above from Mabul shows another similar hybrid, prominently displaying the extensive blue scaling along the gular and pelvic regions, as well as the posterior dorsal blotch. This specimen was photographed in Mabul, Northeastern Borneo. Again, this region is too far north for C. filamentosus to occur, but is well within the distribution ranges of C. tonozukai and C. rubripinnis. While we cannot be certain what parental lineage is involved in creation of this specimen without extensive genetic tests, we can at least hypothesize the species involved here.



Interstingly, these hybrids often lack the filamentous extension of dorsal spines 10 and 11 that would otherwise be present in C. tonozukai. Occasionally, the region is very slightly fitted with a bud, suggesting a very small vestigial filament in place of where it should be in C. tonozukai.
Hybrids of C. filamentosus and C. tonozukai have never been reliably documented, but would likely occur and should be looked for in East Timor where both species overlap. It is also curious as to what this presumed hybrid is hypothesized to look like. Will it possess a long dorsal filament in line with both parents? Sometimes, obvious characteristics are eliminated during hybridization, seeing as genetic recombination is not something we can predict reliably in black and white. If so, will the hybrids look similar to those seen in C. tonozukai x C. rubripinnis?

The specimen above photographed in Bali is one such example where we find discrepancies and difficulty in explanation. The phenotype recalls that of C. tonozukai x C. rubripinnis from Sulawesi and the southern Philippines, but its presence in Bali suggests otherwise. Despite the very obvious hinting of C. rubripinnis in this phenotype, the Lesser Sunda Islands are located too far south for the species to occur. One possible explanation is that C. rubripinnis is able to travel down the Makassar Strait as larval waifs, a channel connecting the Sunda Archipelago and the southern Philippines. The straits receive a current flow of 11 Sv (million m3/s) (Gordon & Susanto, 2006) from Sulawesi to the Banda Arc, making it quite feasible for the species to travel down before winding up in Bali.

Another scenario is that this phenotype is actually a hybrid of C. filamentosus x C. tonozukai. The former species is very common in Bali and the Lesser Sunda Archipelago, while the latter species is more than likely capable of straying here from Sulawesi. If this is so, then the lack of a dorsal filament is very curious indeed, and is in line with the possibility of this feature being hidden and unexpressed during genetic recombination of the two parental species. The biogeography of the species involved makes this a more likely scenario, but, again, without extensive genetic analysis, the exact parental lineage cannot be concluded from just photos alone.
The filamentosus group is far from uninteresting, in this sense, and provides us with another interesting glimpse into the messy speciation which abounds in the Coral Triangle. This discussion should also serve to highlight the difficulties in how best to classify what are often poorly known and heteromorphic fishes. We anticipate the confusion regarding the legitimacy of Cirrhilabrus marinda as a distinct evolutionary lineage (i.e. a “species”) will continue for quite some time.
This article, the 11th in our review of Cirrhilabrus, wraps up the discussion on the long-finned fairy wrasses. Join us next time for the final installment, when we ponder a true oddity in the genus – Cirrhilabrus laboutei – and take a look at what some preliminary genetic data has to say about all this.
