
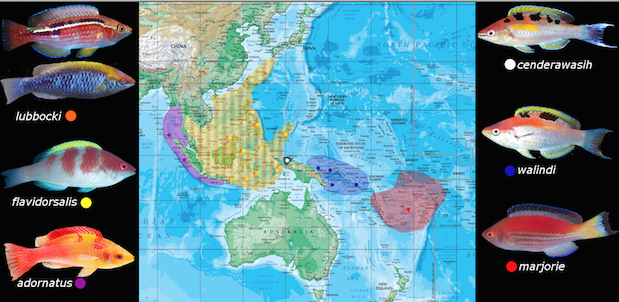
The lubbocki group houses six members divided evenly into two separate but closely related clades. They are the lubbocki clade and the marjorie clade. All members of this group are small to medium sized species with a recurring red and white color scheme in males. Apart from the biogeographic and phenotypic differences of the two clades, the entire lubbocki group share very similar and unique traits that set them apart from the other Cirrhilabrus.
Females of all six species are uniformly pink with a single caudal peduncular spot, and are indistinguishable from each other. Successful identification of females can be carried out based on contextual clues, such as the presence of males or by provision of the geographical range. This is fairly straightforward for four of the members with allopatric and disparate distributions, but gets frightfully confusing in the sympatric C. lubbocki and C. flavidorsalis.

With the exception of C. lubbocki and C. flavirodalis, the remaining four species in this group are allopatric. The overlapping nature of the former throws into question the relationship between the two species, seeing as sympatric speciation is rarely observed in Cirrhilabrus. We will touch more on this later.
In Joe’s previous article, two hypothesized phylogenetic models were presented showing three distinct lineages. The precise relationship of the lubbocki group to other Cirrhilabrus species is poorly understood and awaits more rigorous genetic study. Unique attributes (like head shape and female coloration) suggest it may belong on a basal branch within the genus, but there is also reason to suspect C. laboutei belongs as the basal lineage instead. Compare these competing theories presented in the phylogenetic trees below.

The distinctive fusiform head shape as mentioned briefly above is diagnostic of all members in this group regardless of their varying life stages. The holotype of C. walindi below for example demonstrates this physical attribute very well. A photo of Cirrhilabrus lubbocki in its female form is also featured below, and its unremarkable pattern serves as a template to all six members of the group. The distinctive absence of even the faintest stripes or secondary features in the females are unique and characteristic to the lubbocki group, and this coupled with its head morphology strongly suggests a basal placement in the Cirrhilabrus phylogenetic tree.
It is also worth mentioning that unlike other Cirrhilabrus, the males in this group mostly lack any intricacies in their median fins. Cirrhilabrus walindi and C. cenedrawasih both have spotted anal fins and a metallic iridescence on the caudal, but apart from these two, the other males are rather sombre, possessing at most a simple margin or a few unobtrusive blotches.

The lubbocki clade

The lubbocki clade features three members. They are Cirrhilabrus lubbocki, C. flavidorsalis and C. adornatus. All are frequent offerings to the aquarium trade with C. lubbocki being the most commonly encountered. The species in this group predominantly occupy the Eastern Indian Ocean and the Indo-West Pacific; with C. lubbocki and C. flavidorsalis enjoying a large overlapping range which includes Indonesia, Malaysia, Philippines, Japan and Palau. C. adornatus is restricted to the Eastern Indian Ocean where it can be found in the narrow strip of Sumatra and the Mentawai islands.
The colour patterns amongst the three species are variable and inconsistent, but morphologically they share similar characteristics. The median and ventral fins are simple without any filamentous appendages, and the caudal fin is only very weakly emarginated.
C. lubbocki, C. flavidorsalis and C. adornatus are shallow water wrasses associated closely with loose rubble substrate, especially the last species. It is not uncommon to find C. lubbocki and C. flavidorsalis in large groups, often joined by other species occupying the same niche such as members of Paracheilinus and other sympatric Cirrhilabrus species.
Cirrhilabrus lubbocki

This is a highly variable species with males coming in two forms, each being subjected to wildly volatile patterns and color combinations. In the first form, males have a yellow to red head shield that runs horizontally along the dorsum to include the dorsal fin. The rest of the body varies from pink, to purple, and rarely red, with the scales often edged in metallic blue to form an intricate crosshatch pattern.

The second male form shares basically the same color scheme as the preceding, but instead possesses a pair of horizontal stripes, both tracing the lateral line and running along the dorsum with one disconnected and placed slightly below the other. The striped form tends to be less colorful, and is very often uniformly opalescent silver just below the yellow or red dorsal demarcation. However intermediates of both forms are not uncommon. The species is in general very haphazard and kaleidoscopic in its coloration and patterning.
Morphologically, the median fins and paired fins lack any embellishments. The ventral fins are short and pointed, and the tail is very weakly emarginated. During nuptial display, the median fins turn white and the males develop a lilac band running through the eye and along the body.

Additionally, C. lubbocki shares a similar fright or nocturnal coloration to the other two species, in which it adopts a blotchy pattern featuring transverse bands. In C. flavidorsalis and C. adornatus, the same blotch pattern can be seen in stressed or frightened individuals as well.
The marjorie clade on the other hand does not develop this blotchy fright coloration, and at least in C. marjorie, the entire fish turns red with unobtrusive mottling. More differences between the two clades will be touched upon later on when we explore Cirrhilabrus marjorie and the rest of its clade members.
Cirrhilabrus flavidorsalis

Cirrhilabrus flavidorsalis is another variable species, but nowhere near as fickle as the preceding. The males have a pink to white ground color, with a red head capsule and a variable number of transverse red blotches. The dorsal fin coloration ranges from the namesake yellow, to red or blue.
Phenotypically speaking, C. flavidorsalis resembles C. adornatus more closely with an overall bone white ground coloration and the possession of red transverse blotches. The red is however more developed in C. flavidorsalis, but it is in less erythric individuals that the real similarities between the two can be observed. Its close assosiation with C. lubbocki in the wild however is curious.
As previously mentioned, the sympatric overlap with Cirrhilabrus lubbocki throughout its range raises some red flags. Closely allied species usually follow the allopatric model for speciation, in which a single interbreeding population is broken up into two or more by extrinsic forces such as lowering of sea levels or erection of a land mass. Over time, the isolated populations diverge from the original to eventually form separate species.
Sympatric speciation on the other hand occurs when a single species diverges in the same range, which is not at all common for this genus. A few possible explanations to this phenomenon can be put forth. The first possibility would be that C. flavidorsalis diverged from the sympatric C. lubbocki through sexual selection. In a swath of identical looking females, having a unique nuptial coloration would serve as an advantage for mate identification and selection, and this sexual pressure could catalyze the evolutionary start of C. flavidorsalis.

Another less likely scenario would be that in the past, C. flavidorsalis and C. lubbocki were indeed allopatric, but somewhere along the lines the extrinsic barriers were removed and the two species were able to overlap again. Having already speciated so far along, the two are unable to hybridize back into a single homogenous species and currently the two sisters swim sympatrically over much of their ranges.
The last and least probable scenario would be that the two species are synonymous with the “flavidorsalis” form serving as the third male morph of C. lubbocki. As mentioned in the forward written previously by Joe, without any published research on whether these variants interbreed or what genetic divergence can be found here, how is anyone to know if there is a single heterochromic taxon, three sympatric species or the current accepted model of two species?
With the exception of these overlapping species, the rest of the lubbocki group is rather straightforward.

[Update] The above hybrid photo was added post completion of this article. In a previous hypothesis discussed above, we attributed the sympatry of C. lubbocki and C. flavidorsalis to the removal of a barrier once keeping the two species apart. Having speciated far enough, the reconnection of both species in the same geographical range have now resulted in the inability to hybridize. A photo of C. lubbocki x C. flavidorsalis has been found, suggesting that hybridization between the two species is possible. In the specimen above, traits from C. lubbocki can be seen as the vestigial manifestation of the double lateral stripes.
However, the hypothesis cannot be discounted yet. Both species might still be genetically distinct enough to undo the divergent process, with hybrids occurring as a great rarity. This would explain the scarcity of photographic documentation despite both species being widely sympatric and abundant.
Cirrhilabrus adornatus

Cirrhilabrus adornatus is the final member of the lubbocki clade. In this simply coloured species, the males are uniformly white and possess a variable (usually two), number of red blotches on the anterior dorsum. The dorsal fin is red, and as with the preceding two species, the tail is very weakly emarginated. Young males are often suffused in pink while terminal males are bone white.
C. adornatus is superficially very koi looking, and shares this uncommon trait of being predominantly white with the other members of the lubbocki group.

In nuptial display, Cirrhilabrus adornatus transforms into a uniform yellow-green, with red developing along the anal fin base, caudal peduncle and caudal fin. The anal and dorsal fins turn white, and a red blotch develops on the posterior segment of the latter. The characteristic red blotches on the dorsum turn white during this event. C. adornatus displays more discernible color changes during nuptial display than any other member of this group.

The relationship between C. lubbocki, C. flavidorsalis and C. adornatus is close, to say the least. The placement of these three species in the same clade is not difficult to grasp, and in the following section of this article we will explore why they warrant separation from the second lubbocki group clade which includes the wholly Pacific C. marjorie and the two closely allied species – C. walindi and C. cenderawasih.
[Update] The above image of C. adornatus in nuptial display was added post completion of this article.
The marjorie clade

Like the lubbocki clade, the marjorie clade also features three members. They are Cirrhilabrus marjorie, C. walindi and C. cenderawasih. However unlike the ostentatious abundance of the lubbocki clade members, all the species in this clade are either rare or unobtainable with regards to the aquarium trade. Of the three, only Cirrhilabrus marjorie is collected for the hobby, albeit rarely and very sporadically. Cirrhilabrus walindi has entered the trade with such scarcity and irregularity that the species cannot in anyway be considered “available”, while Cirrhilabrus cenderawasih has yet to make its aquarium debut.
All three species in this group are wholly pacific, with two species having restricted ranges allied to their endemism. Cirrhilabrus cenderawasih is known only from Cenderawasih Bay in West Papua, while Cirrhilabrus marjorie has not yet been discovered outside its type locale of Fiji. Cirrhilabrus walindi enjoyes a slightly broader range, being found in Papua New Guinea to the Solomon Islands.
Phenotypically, the three species are rather similar, being mainly white in base coloration. Cirrhilabrus cenderawasih and C. walindi are extremely similar and are obviously allopatric sisters, while C. marjorie lacks an immediate geminate twin.
Despite the similarities with the lubbocki group, the marjorie group can be distinguished based on a few things. Biogeographically speaking, all the species in this clade are closely allied and are strictly Pacific, unlike the former, which strays into the Eastern Indian Ocean. Morphologically, the marjorie clade members strongly possess double emarginated tails with trailing filaments on the ends of each lobe. They are also fond of deeper waters, and can be found at depths of 160ft and above, in which C. lubbocki, C. flavidorsalis and C. adornatus never reach.
Cirrhilabrus marjorie

Cirrhilabrus marjorie is a relative newcomer to the genus, being described in 2003. It was known from three specimens collected in Fiji, and was named in honor of Marjorie Awai, wife of Dr. Bruce Carlson. In this beautiful species, the males are alabaster with the head and dorsal half a deep crimson, bleeding slightly into the pectoral fin base to form a stripe. The body is marked with four to five horizontal purplish-grey stripes, each occupying a separate scale row. It has a grayish-white dorsal fin with a black margin edged in blue. The caudal fin is yellow with a sub-marginal white bar posteriorly, and a broad black posterior margin.
Cirrhilabrus marjorie has a double emarginated tail which often looks lunate when contracted. This tail morphology is a key diagnostic feature of the members in this clade. C. marjorie is fond of rubble slopes of moderately deep waters where it swims with the likes of Cirrhilabrus punctatus and Paracheilinus rubricaudalis. It has not been documented outside of Fiji where it was first discovered, although it may possibly extend further out.
In its description papers, it has been suggested that C. marjorie is more closely related to C. bathyphilus and C. exquisitus based on the tail morphology. In comparison to C. walindi and C. cenderawasih, the similarities between the three are more believable than what has been originally suggested, and in future installations of this series we will explore that Cirrhilabrus bathyphilus is key to its own group with the allied Cirrhilabrus nahackyi; a species which at the time of C. marjorie has not been described yet.
Cirrhilabrus walindi

Cirrhilabrus walindi is an enigmatic species with very little documented photos. The specific epithet walindi comes from the Walindi Plantation Resort in Kimbe Bay, the type location for the species. In Cirrhilabrus walindi, the male coloration is whitish to pale mauve with a more intensely colored dorsum. The dorsal fin is hyaline yellow with two large black blotches that readily sets this species apart. The caudal fin is doubly emarginated and is a lustrous purple-blue featuring two trailing filaments.

C. walindi is known from Eastern Papua New Guinea to the Solomon Islands where it is fond of waters exceeding 100ft, however it has been reported to swim in depths as shallow as 30ft in Kimbe Bay. Cirrhilabrus walindi has an unusual track record in the aquarium hobby. It appears to have been imported once or twice in the Japanese market many years ago, but has disappeared since. Its scarcity in the trade is rather unusual given that collection in the Solomon Islands is not all that irregular.
Moving to West Papua, it is replaced by an allied sister species – Cirrhilabrus cenderawasih.
Cirrhilabrus cenderawasih

The beautiful Cirrhilabrus cenderawasih is the final member of the marjorie clade as well as the lubbocki group in general. This species is most closely related to Cirrhilabrus walindi and the two species likely arose from the same ancestral species that once swam the margins of the New Guinea-Australian plate. Within the past few million years, opportunities for speciation must have arose which gave rise to allopatric members in the Cenderawasih region.
A series of tectonic processes have been responsible for shaping the region’s geology over the past fifty million years. According to Hill and Hall’s (2003) paleoreconstruction model, a series of island arc terranes arose along the edge of the Caroline Plate, eventually colliding with the Australian Plate, and then sliding parallel to the northern coast of New Guinea before eventually accreting to form the northern ranges.
One of these arc terranes known as the Tosem Block (Hill and Hall, 2003) slid across the entrance of Cenderawasih Bay between two to five million years ago, before finally stopping just at the northern edge of the Bird’s Head Peninsula. This isolated the region, restricting movement into and out of the bay and setting the stage for allopatric speciation. The shifting of plates work in tandem with the region’s fluctuating sea levels, further amplifying the allopatric isolation and promoting the rich development of unique species within the Cenderawasih bay area.
C. cenderawasih shares the same basal coloration as the preceding species, but instead has a strong and characteristic yellow rectangular blotch smeared across the middle of its body. The stripe only reaches halfway. Above, four to five irregular black blotches pepper the dorsum. The blotches are highly variable in size, and very often the first one is vestigial or even completely absent.

C. cenderawasih is common at depths exceeding 100ft, where they swim in aggregations often surpassing twenty individuals with females greatly outnumbering males. In the holotype specimen above, the filamentous tail is very clearly seen. The central caudal rays are longer, giving the leading margins on either side of it a concave appearance leading to the characteristic double emarginated tail.
With the three species of the marjorie clade elaborated and touched upon, it’s easy to see the resemblance amongst each other. From an evolutionary standpoint, C. walindi and C. cenderawasih are immediately related, with C. marjorie being rather closely grouped alongside based on morphology and biogeographical distribution.
It is easy to see why the lubbocki and marjorie clades are not merged together, based on difference in classified range, morphology and habitat preference. However the two clades clearly share a similar relationship especially with regards to the nearly identical females of all six species.
The lubbocki group is a fascinating start to this long and challenging series, and offers a glimpse into various recurring themes that we will explore such as species groupings, biogeography as well as evolutionary models of speciation. In addition, it is one of the rare instances where two members of the same group are sympatric. In future installations the species groups will only become more complex and confounded, but we hope you’ll stick around until the end to fully soak in the rich diversity that this genus has to offer.
