

Known for their dazzling colors, swaying tentacles, and propensity to sting nearby corals, the Torch Coral, Euphyllia glabrescens, has been a staple of the reef aquarium hobby for decades. And given the continuous discovery of new varieties, this species will likely retain its status for many years to come. However, while Torch Corals, or “Torches” for short, have become nothing less than a household name among reefers, many of the details surrounding their history, taxonomy, and biology often go overlooked.


What we know now as Euphyllia glabrescens was first described in 1812 by Adelbertus de Chamisso and Carolus Guilelmus Eysenhardt as Caryophyllia glabrescens. The original description translated (roughly) from Latin is as follows “Dichotomous, from here to trichotomous. Branches as thick as a finger, or thicker, glabrous on the outside or obsoletely striated, the trunk glabrous. The star is hollow, with a very deep center, and the lamellae are entire or obsoletely toothed, alternately larger.” The authors named this new species “glabrescens” after its glabrous branches and trunk, owing to the relatively smooth exterior of the coral’s skeleton. Many other Euphyllia and Fimbriaphyllia skeletons bear raised external striations that run down the length of their branches.
In 1846, an American geologist named James Dwight Dana erected a new genus, Euphyllia, based on broad skeletal similarities among stony coral species that were known at the time. The type species for Dana’s new genus was none other than Euphyllia glabrescens. Many other scleractinian taxa unified by having large corallites with smooth septa would at some point be seated within the genus Euphyllia. Eventually, the majority of these species including the entire genus Fimbriaphyllia, Catalaphyllia jardinei, Plerogyra sinuosa, and Truncatoflabellum spheniscus would be reclassified into completely unrelated genera as the scientific community’s collective understanding of coral systematics improved. However, despite the taxonomic turmoil experienced by other members of its genus, E. glabrescens is an epithet that has withstood the test of time and remained unchanged for nearly two hundred years.

The modern definition of what exactly qualifies as Euphyllia glabrescens has thankfully been refined over the past two centuries. E. glabrescens is described as having phaceloid coralla that are taller than that of its congener E. cristata. Corallites attain a size between approximately 20 and 30 millimeters in diameter, with the space between individual corallites generally ranging between 15 and 30 millimeters depending on environmental conditions. Within each corallite, primary and secondary septa plunge deeply toward the interior. Each septum is smooth, lacks paliform lobes, and is not exsert. Like other euphyllids, the corallites of E. glabrescens lack columellae at their innermost portions.
However, unlike many other stony corals where isolated skeletal elements are the most critical component lending to proper identification, many euphyllid corals like E. glabrescens can be only properly diagnosed when their living polyps and skeletons are both taken into consideration. E. glabrescens polyps have long, tubular tentacles arranged in multiple concentric cycles, not entirely unlike that of a sea anemone. Each tentacle terminates with globular or sometimes ovate acrospheres that are a distinct color from the rest of the polyp. These tentacles may feature abnormalities including bifurcation, a crinkled or swirled appearance, improperly positioned acrospheres, and asymmetrical protrusions of varying shapes and sizes.
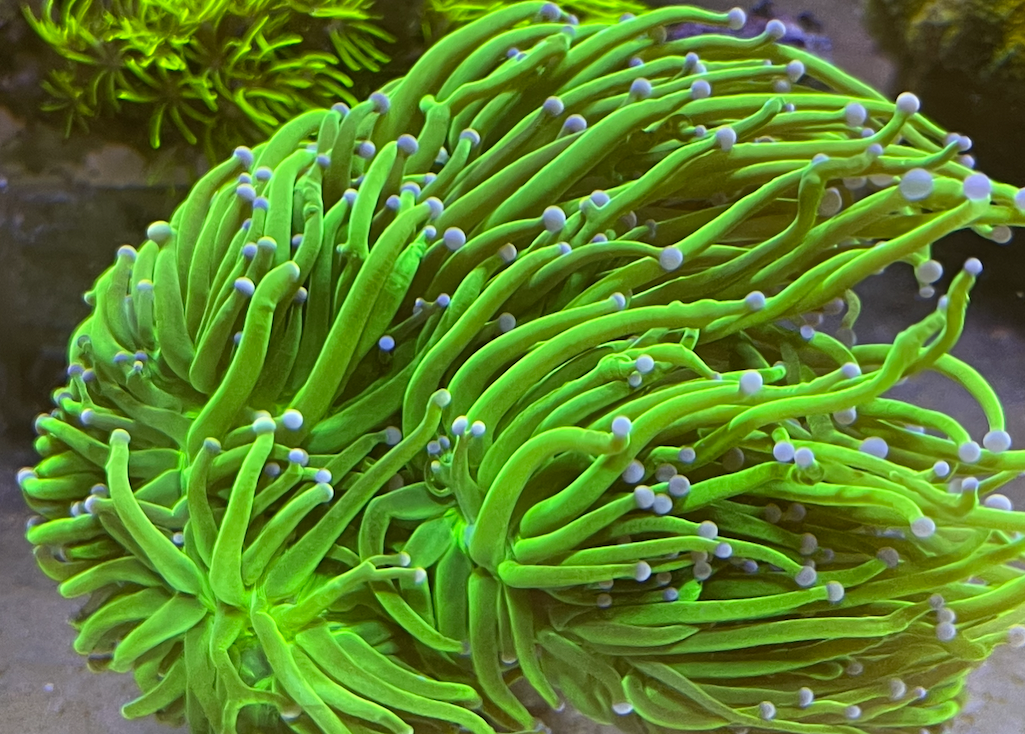


There are apparently some nuanced differences between Australian and Indonesian Euphyllia glabrescens as well. Many avid Torch Coral collectors note that Australian varieties exhibit thicker, more cylindrical tentacles, while their Indonesian counterparts have tentacles that are much thinner and taper distally toward the acrosphere. Beyond physical appearance, many aquarists seem to have experienced slower growth rates or outright lesser success with Australian Euphyllia glabrescens over Indonesian ones. The underlying cause(s) of these differences remains unclear.

The geographic distribution of Euphyllia glabrescens spans the entirety of the broader Indo-Pacific region and is even reported from the Red Sea. In situ, Torch Corals are a conspicuous, but relatively uncommon sight on the reef. While this species can be found in a variety of habitats they tend to be most common in areas with sheltered waters ranging from the intertidal in some locales to a depth of around 100 meters. Torch Corals usually occur singularly, with just one colony in any given spot, occasionally surrounded by smaller colonies that are presumably offspring or detached fragments that have since settled. These colonies typically attain a hemispherical shape, approximately a foot in diameter, but much more expansive colonies have been reported.

While the history surrounding the cultivation of Torches in captivity is spotty at best, we know that aquarists have kept Euphyllia glabrescens in aquaria since at least the early 1990s and would not be surprised if the earliest successes of captive E. glabrescens predated this by at least another decade or two. Since the 2010s, the reef aquarium hobby has seen a large push toward aquacultured Euphyllia, although imported maricultured pieces still make up a percentage of the Torch Corals in circulation.
Maricultured corals present somewhat of a double-edged sword. On one hand, they have the potential to increase the diversity of Torch Coral varieties that exist within the hobby. However, these corals often come in with a suite of invertebrate and microorganismal hitchhikers with varying degrees of desirability. In a worst-case scenario, Euphyllia-eating Flatworms (EEFW) may stow away on wild or maricultured Torch Coral colonies. These flatworms voraciously consume the coenosarc at the base of E. glabrescens polyps, causing the coral to retract and slowly decline in health. If left untreated the coral will eventually succumb to the infestation. Fleshy “Large Polyp Stony” (LPS) corals like E. glabrescens are also susceptible to polymicrobial infections. Collectively known as Brown Jelly Disease (BJD), these infections can spread rapidly from one corallite to another, and even jump between colonies in a matter of days. To date, the treatment of BJD has been mostly relegated to damage control, but proper husbandry practices and new techniques like polycropping can help to mitigate the risk of a BJD outbreak.

Luckily, Euphyllia glabrescens is more than capable of asexual reproduction, whether through manual propagation (i.e. fragging) or of its own accord. Cutting through the non-living skeletal portion of the coral allows for the easy division of the colony. Whether this is conducted, to sacrifice a polyp infected by BJD or to create frags for a friend, E. glabrescens is one of the easier corals to propagate. If left alone, E. glabrescens will grow via a process known as intratentacular budding, colloquially known as “splitting”. During intratentacular budding, (see skeletons images above,) a single polyp will elongate laterally before diving into two polyps or near identical size and shape. Mature colonies of E. glabrescens may also form asexual buds at the base of their polyps, but this is less common than in Fimbriaphyllia spp.


E. glabrescens has also been noted to sometimes exhibit polyp bailout, where the entire polyp detaches from its corallite and floats into the water column. Typically this occurs under adverse conditions and is typically seen as a “last-ditch” effort toward survival, however, this is purely anecdotal. In some instances, detached polyps may settle in low-flow areas and begin to reform their skeletons. However, survival of these bailed-out polyps is incredibly rare, even with intervention by the aquarist.
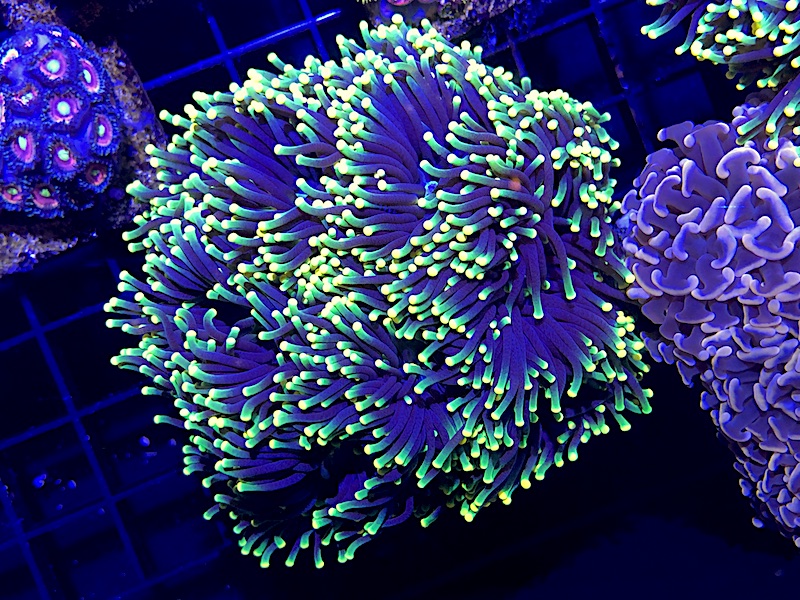
Despite all the struggles associated with the husbandry of Euphyllia glabrescens, we continue to see promising breakthroughs in the industry. New coral dips seem to have encouraging results in effectively eradicating adult EEFW and treating BJD. Additionally, the hobby has seen marked success in the captive spawning of Euphyllia glabrescens and the subsequent settlement and cultivation of these sexually-reproduced corals. What makes E. glabrescens such a great candidate for this is their mode of reproduction. While many stony corals are gonochoristic (having separate male and female sexes), Torches Corals are hermaphroditic, being able to produce both sperm and eggs from a single colony. Additionally, while the vast majority of stony corals are broadcast spawners, E. glabrescens fertilizes its eggs internally, brooding the planulae (coral larvae) inside their polyps.
Ultimately, this results in fewer but more well-developed planulae that may potentially possess an edge over their broadcast-spawned counterparts as recruits. The concept of sexually reproduced corals is particularly exciting because of the increased genetic variation within spawned progeny due to homologous recombination, independent assortment, and random fertilization that occur during meiosis and fertilization. This in turn provides the potential for near-infinite genetic combinations which could result in a variety of desirable traits ranging from new color combinations to varieties that are more resistant to bacterial infection.

Clearly, Torches are more than just a swirling mass of colorful tentacles. With so many questions left unanswered, I am excited to see what’s in store for this iconic stony coral species, which presents a burgeoning frontier for coral research and the aquarium hobby alike. Perhaps future taxonomic studies will uncover cryptic genetic diversity between Australian and Indonesian Euphyllia. Maybe, we as hobbyists will develop ways to selectively breed captive Euphyllia to maximize the expression of desirable traits.
About the author
Sean Ono has been keeping corals for the better part of a decade, both at home and as a former volunteer aquarist at one of the largest public aquariums in the United States. He has a particular affinity for anthozoan cnidarians (including corals) but is passionate about echinoderms, marine worms, macroalgae, and more. Despite his familiarity with reef-associated organisms, his previous academic research is more elasmobranch-centric, aiming to quantify the biomechanical and behavioral aspects of stingray tail strikes. Having recently graduated with a Bachelor of Science in Marine Biology, Sean hopes to pursue a Ph.D. in ecology and evolution, focusing on the intersection of biomechanics, sensory capabilities, and comparative morphologies of marine taxa.
