

On social media, I often see the question, “Why is my coral dying?” In this article, I will explore how complicated it is to answer that question even from a scientific standpoint.
In previous articles, I have covered the idea of the holobiont, coral immune system, and how pathogenic events occur through a system of dysbiosis instead of the one pathogen, disease model. Here, I will synthesize a much more complete view of the molecular pathways that interact with each other to cause coral disease – my goal is to illustrate just how complicated all of this is and hopefully generate a healthy amount of skepticism toward anyone who claims to have a “solution” to coral disease without sufficient evidence and explanation of a mechanism that can interact with these systems.
It Started with a Whisper
When corals are healthy, they exist in relative stability, with a conserved density of symbiodinium, microbial community, and metabolic rates. While some of these processes ebb and flow with the seasons, tides, and depth on the reef, most coral species are now believed to have reasonably consistent core “omes” (whether it be microbiome, metabolome, or proteome). These are thought to exist in a gradient among species on a reef with small shifts due to spatial dispersion. Due to the stability of the ocean and the fact that corals have existed for millions of years, continually adapting to that environment, the sudden intense abiotic swings they are now experiencing due to climate change are entirely out of the ordinary and are the root cause of many of the issues now observed on reefs. Decades of research analyzing coral from various angles have answered many questions about what occurs when a coral encounters abiotic stress. It is widely supported that external stress (biotic or abiotic) is the lynchpin that causes corals to decline, whether from bleaching, disease, or both. Several distinct shifts occur once a coral is exposed to stress.
They Came from Within
Typically, the association between symbiodinium and the coral begins to weaken as this swing hits the zooxanthellae hard. This results in less productive photosynthesis and the activation of several energetically costly metabolic stress relief pathways, such as the production of heat shock proteins, catalases, and phenoloxidases. Upregulating these alternative pathways leads to excess dimethylsulfonoproprination or DMSP (more on this later). Holistically, this leads to corals receiving much less energy from photosynthesis. This leads to an upregulation of fatty acid metabolism, which momentarily allows the coral to access an alternative energy source as it releases its stored carbon supplies. The breakdown and death of zooxanthellae cells, combined with excess carbon released from within the coral tissue, leads to a significant shift in the coral mucus layer’s fatty acid, amino acid, and protein composition. Typical metabolism accumulates a small number of carbon sources in the mucus. Beneficial bacterial symbionts selectively metabolize these carbon sources. When the chemical composition of the mucus shifts, the microbiome is correspondingly affected.
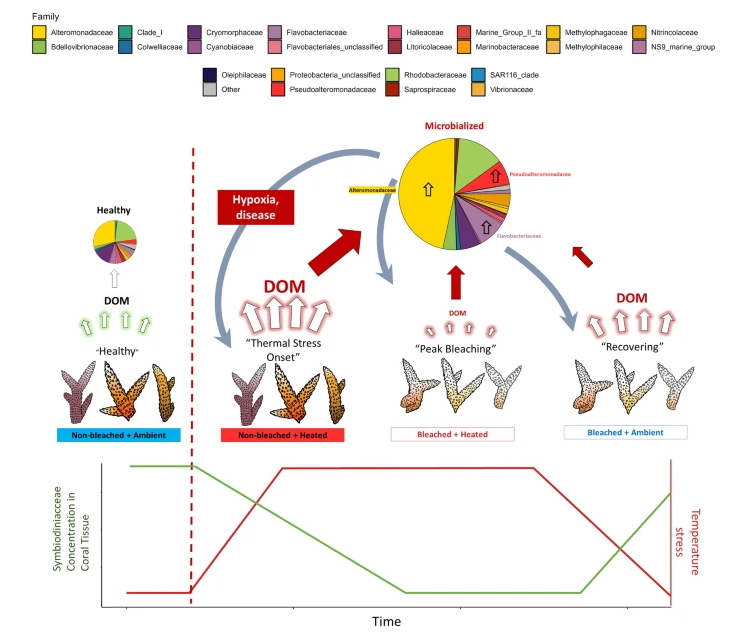
Scheme illustrating connections between mucus composition and the “microbialization” of the microbiome during pathogen events https://www.nature.com/articles/s42003-023-05730-0
It’s important to understand that the bacteria that make up the coral’s microbiome each serve a different function. Some of these species are purely “good guys” – these species, such as Endozoicomonas, act as central regulators of the microbiome where they break down various waste products from the coral and zooxanthellae and turn them into compounds designed to alter the gene expression and population growth of other species in the holobiont. Some species work as “contractors” and can be described as “frenemies.” These species are typically regulated very heavily and kept at trim population levels by the good guys due to their potential to quickly grow, express virulence factors, and ultimately overtake the coral if presented with the opportunity. Many of these species are important in nutrient cycling – some examples are bacteria in the family Rhodobacteraceae, Cyanobacteria, and Flavobacteriaceae. These species are “hired” by the coral to perform dirty jobs, but if allowed better pay, they would quickly turn their back on the community they work in. Thus, the microbiome is in a constant balancing act that’s on very thin ice. These frenemy bacteria become more competitive whenever an abiotic shift occurs as a larger diversity of carbon sources becomes available. Growing evidence shows that many “good” bacteria can only metabolize the few carbon sources healthy corals typically produce and cannot correspondingly metabolize other carbon sources.
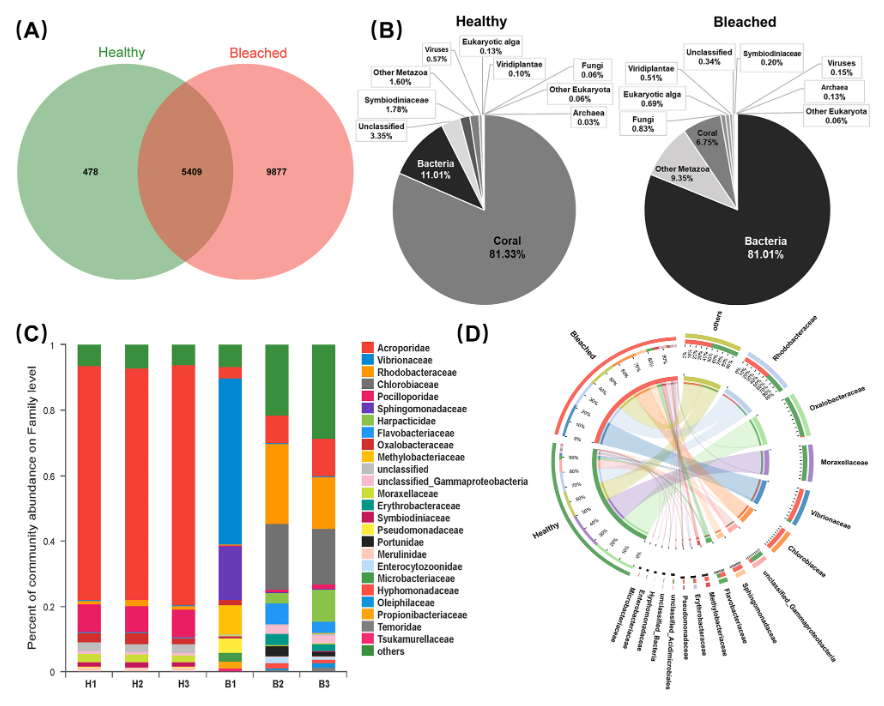
An illustration of how bacterial taxa shift from healthy to bleached corals https://journals.asm.org/doi/10.1128/msystems.00505-23
In contrast, some bacterial symbionts that exist in smaller concentrations have the genetic potential to metabolize an extensive array of carbon sources. Two phenomena occur as these species continue to grow and outcompete the standard mutualistic symbionts. First, as the “good guys” die, the critical regulatory compounds they produce, which help control the population and gene expression of the frenemy bacteria, decrease concentration. Second, the waste compounds the good guys eat begin to increase in concentration. These frenemy bacteria take on an r-strategy where they opportunistically take advantage of excess resources in a chaotic environment and seek to grow as fast as possible. This shift in metabolite concentration compounds the internal dysbiotic event as the “revoluting” bacterial taxa grow faster and faster as the concentration of inhibitory compounds drops off and the build-up of external info chemicals that the holobiont would typically break down begins to build in concentration, creating another facet of this problem…
Moths to a Flame
As good bacteria continue to die, important infochemicals such as DMSP typically quickly break down and rapidly build up in the coral mucus and surrounding water. Many external coral pathogens have evolved to detect this chemical. In Vibrio, once DMSP is detected, many genes are activated, initiating virulence factors. Toxin production, secretion systems, biofilm formation, and rapid growth genes are then turned on as Vibrio swim directly towards high concentrations of DMSP and find the stressed corals like moths to a flame. Once they’re at the coral, they express glycoside hydrolase genes that allow them to break through the thick peptidoglycan concentration of the coral mucus and successfully adhesive to coral cells. Typically, multiple external pathogens will chemotax to and infect the coral. New evidence illustrates that when multiple strains of Vibrio infect a coral, the virulence factors they produce synergistically work together, leading to a much more aggressive response.
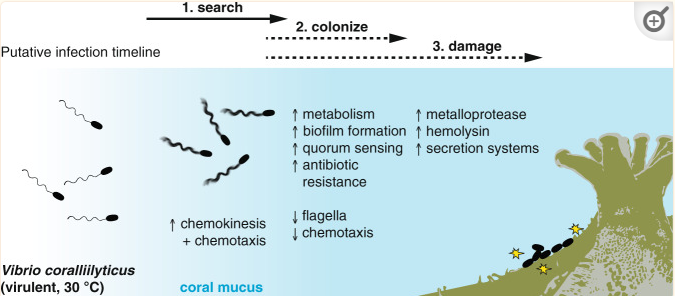
Pathway for Vibrio infection https://www.ncbi.nlm.nih.gov/pmc/articles/PMC8630044/
Additionally, through competition sensing, the two Vibrio strains produced external virulence factors, which led to differential expression in the opportunistic “frenemy bacteria,” which allowed them to shift to a purely pathogenic state, further accelerating the already ongoing r-strategy pathogenic pathway of the internal opportunistic are enacting. In response to this stress, corals produce Lysa-PAFs (platelet activating factors), which cause the upregulation of ROS (reactive oxygen species). This is a last-ditch attempt that indiscriminately damages the coral and pathogenic bacteria alike. This directly led to host tissue damage and necrosis, further pushing the coral to the edge.
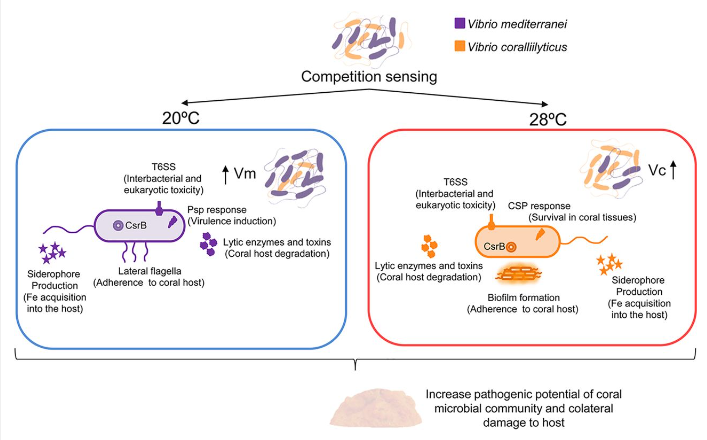
Vibrio competition sensing mechanisms https://journals.asm.org/doi/10.1128/mbio.00201-20
Further Complicating Things
On top of this, new evidence suggests that both symbiotic and pathogenic external bacteria directly regulate the coral’s gene expression in the instance of pathogens; it is believed that they may be able to produce exomes released through their virulently expressed secretion systems that contain ankyrin-repeat proteins (ARPs). In particular, the protein AnkA has been shown to move into the nucleus and bind to portions of chromatin that control the expression of genes that lead to ROS production in human pathogenic bacteria. Many coral and sponge-associated bacteria are now proven to contain large amounts of ARPs; thus, some of this functionality may be conserved. If this hypothesis is true, that would mean that when a coral is infected by a bacteria that has the genetic potential to form a secretion system as well as ARPs, that bacteria may be inducing epigenetic shifts in the coral, which leads to the downregulation of the coral’s innate immune system. This makes the issue of coral disease even more complicated as approaches that aim at upregulating the coral’s native immune response may be futile without first addressing this complex mechanism.
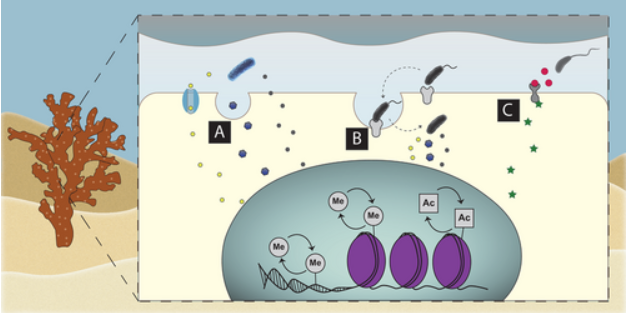
Speculative pathway for how epigenetic regulation may occur between bacteria and coral. https://onlinelibrary.wiley.com/doi/full/10.1002/bies.202100068
Putting Everything Together
Thus, in as simple a pathway as I can provide from current literature, your coral is likely dying due to the following chain of events.
- Abiotic or biotic stress
- That causes zooxanthellae to become stressed, which leads to less efficient photosynthesis, which leads to the coral-eating carbon sources it saves up.
- That leads to the carbon sources shifting in the mucus, allowing opportunistic bacteria to outcompete good core microbes.
- That leads to the build-up of info chemicals that recruit external pathogens.
- Those pathogens then directly shut down the innate immune system by regulating the coral’s genetic expression and upregulating the virulence of the localized opportunistic bacteria that have already started to take over the coral from within.
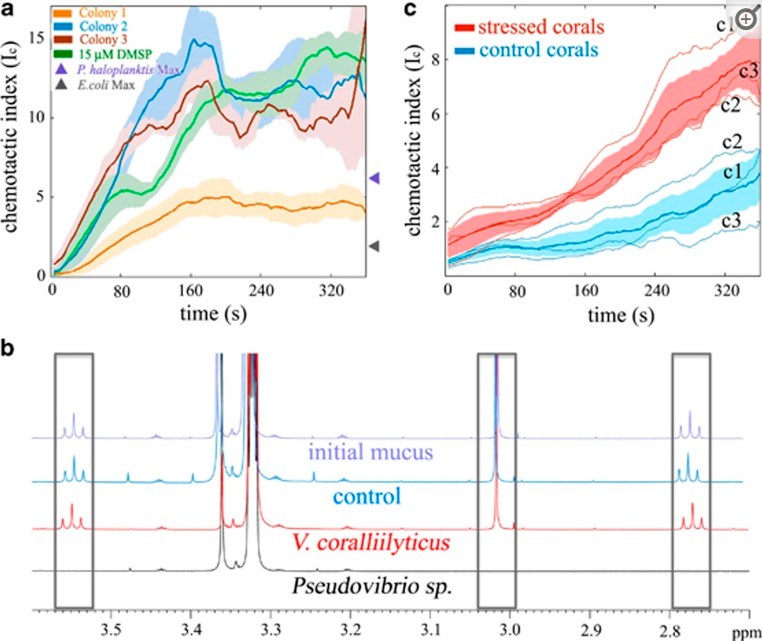
Data illustrating DMSP as the primary info chemical responsible for Vibrio chemotaxis https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3996689/
What makes this even more complicated is that this chain of events can occur with a consortium of different bacterial species. While the overall function of the bacteria may be the same across species (regulatory species, opportunistic species, or pathogen), a confluence of species can fill these roles. Simply put, “This phenomenon concurs with the Anna Karenina Principle (AKP) in the microbial field (34, 35), which states that “all healthy microbiota are alike; each disease-associated microbiota is sick in its own way.” (Cheng, 2023). What makes coral disease a hard problem to conserve is that the disease state manifests distinctly depending on the function of the local environment and its spatiality. Thus, the typical one-size-fits-all strategies of modern medicine are unlikely to solve an issue we still know little about after nearly 40 years of international research. To solve a novel problem – we need a novel solution. Hopefully, in the coming years, one will rear its head.
Sources:
https://journals.asm.org/doi/10.1128/msystems.00505-23
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6497039
https://journals.asm.org/doi/10.1128/mbio.00201-20
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6925168
https://www.int-res.com/abstracts/meps/v322/p1-14
https://www.nature.com/articles/ismej20169
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4805648/pdf/fmicb-07-00371.pdf
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC8630044
https://onlinelibrary.wiley.com/doi/full/10.1002/bies.202100068
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6497039
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3996689
https://www.frontiersin.org/articles/10.3389/fmars.2022.912862/full
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6040183/pdf/e00791-17.pdf
