
The temminckii group plays host to some of the genus’ larger and more unappealing species. Burley, pugnacious and often glazed in matte grey, these un-fairy looking fairy wrasses are more often than not associated with the ugly duckling sobriquet. Despite their lackluster charm and uninspired appearance, the temminckii group members display a textbook example of speciation in its incipient stages, as well as the inconsistencies in the treatment of various taxa across the board.

Cirrhilabrus temminckii has a rivaled history with Cirrhilabrus cyanopleura in being the earliest scientifically recognized Cirrhilabrus. Although technically Cirrhilabrus cyanopleura was discovered first by Bleeker in the early 18th century, its placement in the genus Cheilinoides left an open space for Cirrhilabrus temminckii to fill. The latter was described in the same year of 1853, making it the first official member of the new genus Cirrhilabrus. It subsequently became the type representative for the genus, setting the morphological benchmark in which all future species would be compared against. C. cyanopleura along with C. solorensis were later placed in Cirrhilabrus as the second and third species, making Cheilinoides superfluous and defunct.
The erection of this new genus set off a Cirrhilabrus rat race, with nearly 40 species described by the end of the 19th century. However, it was only 35 years later since the discovery of Cirrhilabrus temminckii did another member of similar stature was described. Cirrhilabrus balteatus became the 19th member discovered in 1988, followed by C. punctatus, C. katherinae and finally C. beauperryi in 1989, 1992 and 2008, respectively.
Despite the advancements in this group over the years, the phylogenetic relationship between its members remains largely unresolved and messy. Both taxonomic and biogeographic distinctions within this group are similarly shrouded in a miasmic haze of ambiguity. This can be seen in the temminckii species complex, where at least four distinct phenotypes can be found distributed from Japan, south to Indonesia, the Philippines and Western Australia. Confusion reigns yet again in the balteatus and katherinae sisters, who, despite nary any phenotypic differences, are awarded with full species status. To further complicate things, C. katherinae has a complex history that may suggest a hybrid origin story. This will be discussed in detail later.
And so, this presents us with some of the most inconsistent, intractable and confusing taxonomic treatments anywhere in Cirrhilabrus, as well as a biogeographic overlay that suggests a smooth, continued transition in phenotype from Cirrhilabrus balteatus to Cirrhilabrus temminckii. This continuum in phenotype gradation could represent incipient stages of evolution, or a complex series of hybrids.
The punctatus and beauperryi sisters in the Melanesian and Papuan regions are slightly less complicated, but are not without their own set of questions. To put things simply, the temminckii group presents the reader with a biogeographic mosh pit of highly similar phenotypes with overlapping ranges and frequent hybridizations. Due to the overt complication in the temminckii and katherinae species flock, great caution must be exercised in discerning the various phenotypes and how they intertwine.
Trying to understand how the complex relates on a phylogenetic level, is to be quite honest, a stretch. In this article, we present the reader with a collection of photos from various locales, and apart from drawing very rudimentary biogeographic distributions and relatedness, much of it is left to be unanswered.

Members of this group can be diagnosed easily based on a set of key characteristics. As with keeping in this lineage, males of all species possess exceedingly long pelvic fins, which always trail past the anal fin origin. The length of these fins rival those of the pylei sisters in the rubrimarginatus group, but are more pliable and filamentous, especially toward the end. Their caudal fins are rounded, often heavily spotted and possesses a blue or golden reflective sheen when viewed at various incidence of light. The caudal scintilla and pelvic fin traits are analogous to all members in the long-finned clade, being seen in the rubrimarginatus, rubriventralis and filamentosus groups.
With the exception of Cirrhilabrus temminckii, all other members lack definition in the dorsal, facial and medial stripes. These are often present as either vestigial or unobtrusive manifestations. In all species, however, the stripes are never solid, but are always either broken, or highly perforated. The temminckii group can be further divided into three clades, each with their own characteristic set of features unique to the members within.

The balteatus clade members are easily recognized in having a light orange girdle on the posterior portion of the body, just behind the operculum. This feature is very strongly displayed in terminal males, but even in initial phased individuals and transitioning females this trait can be identified with scrutiny. During nuptial display, this girdle lightens to white and disappears momentarily. The dorsal and anal fins also possess prominent medial stripes, which are often outlined in blue. This medial stripe pattern is a salient feature, being found in all members of this group except the temminckii complex. Only one phenotype there has this feature, and it is restricted to Cebu.
Members of the punctatus clade lack any such embellishments, but, instead, possess a heavy dusting of metallic spots all over the body and fins, as well as lacking a prominent girdle. This characteristic spotting is translated even in females and immature males, making identification very easy. Together, these two clades are easily separable from the temminckii clade by their lack of dorsal and facial stripes.

Females and juveniles of this group are rather generic looking, uniformly orangey with a white belly, and possess a single black spot on the caudal peduncle and a snout tipped in white. Females from the punctatus clade often possess minute spotting over their entire body, but those from the other clades are often confused with females of C. exquisitus, and cyanopleura. As such, they make for rather unreliable diagnostics.
The temminckii clade

The temminckii complex represents a species flock with at least four or more distinct phenotypes spread across the Western Indo-Pacific, from Japan, south to the Philippines, Indonesia and Western Australia. These varying phenotypes form a spectrum of variation from north to south. The Japanese population of Cirrhilabrus temminckii is an important species, serving as the type specimen for its complex, as well as the type species for the entire genus.
Cirrhilabrus temminckii (Type)

This phenotype is the type representation for the species, as well as the entire genus. As such, all geographical variants or potential cryptic species should be referenced against this. Cirrhilabrus temminckii is a medium to large fish. In this species, the ground coloration is a dingy yellow to beige. The entire dorsum is orange red, spreading into the cranial facial region but ending just along the lower orbital limit, where it is delimited by the distal eye stripe. The reddish back fades ventrally into a yellow-orange suffusion, before intensifying into moss green medially along the body. The extensive red back is a fairly diagnostic feature of the type temminckii, with the other complex members either lacking it entirely, or has it much reduced.
A pair of facial stripes are present, with each situated along the upper and lower orbital limits. The upper stripe extends horizontally to form a complete and fully intact dorsal stripe. The lower stripe extends to form a medial stripe, but this is always incomplete and variably restricted to the anterior preopercular and posterior caudal peduncular region. This stripe is never connected in-between. All the usual body stripes are metallic blue and perforated.
The dorsal, anal, and caudal fins are cellophane bluish to purple. In C. temminckii, the dorsal and anal fins are always without a true medial band. In some specimens, a dark suffusion may be present along the entire dorsal fin length, but this is always unbordered by blue and indistinct.


The species possesses a rather prominent caudal scintilla, which, at various incidence of light, takes on a highly reflective metallic sheen with foil like qualities. This feature is likely analogous to the other groups in this lineage.



C. temminckii takes on a remarkable change in appearance during its nuptial display. In its excited state, the body coloration darkens, while all the usual body stripes scintillate in an electrifying cerulean. It is in this state that the posterior medial stripe becomes more clearly seen. The caudal scintilla during this event becomes extremely chatoyant and reflective. The species is also able to puff up its gular flap, which it extends prominently during acts of aggression or nuptial display. Like the rubrimarginatus group, the males are able to independently control the movement of each pelvic fin, twitching them like a pair of chopsticks.
In full nuptial display, the entire body lightens to a pale, downy white. The dorsal stripe may be so intensely electrifying that it extends in brightness throughout the entire dorsum. This is a trait exclusive to the type species of C. temminckii, and is useful in separating the various similar phenotypes. The upper and lower eye stripes glow in a similar manner, and at the height of nuptial display, the two ends of the broken medial stripe may momentarily appear to connect (although this rarely happens).
The resting and nuptial coloration of this species are salient features to consider before moving on with this article. The various phenotypes within this species complex will be compared against this type specimen, and the minute differences in coloration will serve to set the various forms apart.


The true Cirrhilabrus temminckii is rarely encountered in the trade. It is infrequently imported from Okinawa and various other Japanese islands. It spreads south to the Philippine and Japan, and although uncommon here, it mixes with a different phenotype. Despite its uncommonness, the fish is rarely expensive and somewhat unpopular. It does not hold its coloration well in captivity, and is reported to be belligerent and bossy.
Cirrhilabrus cf. temminckii (Philippines – Cebu)

This next phenotype is instantly recognizable. This highly variable form is orangey-red, to terracotta, to gunmetal grey or green on its dorsum. The ventral region is white to light magnesium gray, and is delimited abruptly by a nascent medial stripe, which, like in C. temminckii, is always incomplete and restricted to the anterior preoperculum and posterior caudal peduncular regions.
This phenotype is characterized by the presence a thick, copious equatorial black belt that runs just above the medial stripe. This belt is segmented and broken into two, with a short anterior section above the pectoral fin base, followed by an equally sized section absent this dark band and a much longer L-shaped continuation of the black belt, which extends through the caudal peduncle. This belt is distinct from the base ventromedial coloration, and are two separate patterns.
The dorsal stripe in this phenotype is extremely vestigial, and not noticeable in the resting phase. This applies to its paired eye stripes as well as its incomplete medial stripe. During nuptial display, these scintillate in blue, but are otherwise heavily obfuscated at rest. This is the only temminckii complex member to lack a noticeable dorsal stripe.
The dorsal and anal fins are dark orange to red, and are strongly marked with a transverse medial band. The band is black, and margined on both sides in blue. Again, this is the only phenotype in the temminckii complex to possess this medial stripe on the dorsal and anal fins, and in this regard, together with the absence of a dorsal stripe, is extremely similar to balteatus and the katherinae complex members. Interestingly, Palau is geographically approximate, and balteatus-like fish occur there. Whether or not waifs into Cebu occur with any regularity is unclear, but it seems plausible. Could this inference mean anything with respect to this phenotype? We’re not certain as of yet.






During nuptial display, the belly whitens to a cotton white while the dorsum darkens to a dusky grey. The proximal half of the transverse medial band disappears, while the rotated “L” distal portion intensifies to a deep black. In nuptial colors, the tail end of the “L” extends ventrally to touch the anal fin base. The usual eye, dorsal and medial stripes scintillate in blue, but these are very weakly expressed even during nuptial flashing.



Excited males may puff up their gular flap and erect the front two thirds of the dorsal fin. The remaining soft portion is pressed against its body. This unusual display is restricted only to members of the long finned clades, and is not seen in any other members of the short finned clades (e.g. jordani, lubbocki, lunatus etc). Cirrhilabrus marinda, C. punctatus and C. rubrimarginatus are also well known for displaying this unusual behavior. In full display, this phenotype whitens considerably to include the basal region of the dorsal fin.
This variant is restricted to the Philippines, more specifically the central region of Cebu. It has not been documented outside of this region, and in the rest of the Philippines and Indonesia, it is replaced by a different phenotype.
It shows up in the trade semi-regularly, but only from Cebuan exports. Like C. temminckii, this phenotype can be quite pugnacious and does not appear to hold its coloration well.
Cirrhilabrus cf. temminckii (Philippines and Indonesia)

This phenotype resembles more closely to the type species in Japan than the unnamed variants in the Philippines. The dorsum, however, is highly variable, ranging from red, to khaki and any shade in-between. It is, however, most often olive-green. The belly is off white to pale gray, and a fuliginous moss green streak is present medially, emerging from the pectoral fin base and variably extending outwards. This streak is highly variable, and can be very extensive, or totally absent.

Like C. temminckii, this phenotype possesses eye stripes on the upper and lower orbital limits. The dorsal stripe is prominent and complete, while the medial stripe is inconsistently developed. In some specimens, this medial stripe is complete and well defined; while in others it remains vestigial and restricted only to the caudal peduncle. The dorsal and anal fins are light brown to red and lack the transverse medial band present in the Philippine phenotype.

Some specimens display vestigial darkened blotches on the body, which recall those seen in the Cebuan cf. temminckii phenotype. What kind of relationship this similarity suggests is unknown, but it appears to be variable, ranging from weakly present to completely absent. One trait that is consistent and separate from the Cebuan population, however, is the complete and prominent dorsal stripe.



In nuptial display, the ventral region lightens to a dingy white-grey and the dorsum darkens to a red or deep olive. The body stripes scintillate in an iridescent blue, lending the apt common name “Blue-striped Fairy Wrasse” to be rather apt. Unlike the Japanese type, the scintillating dorsal stripes are very well defined and neat, and the dorsum appears unable to lighten to a bright white.

As previously mentioned, large males may variably possess darkened blotches on the body and pectoral fin bases. Whether or not these are indicative of something more is unclear, as various specimens with and without these markings have been documented throughout its range. During nuptial display, these may intensify to form a medial band as seen above. A white streak also develops, covering the underlying fuliginous green streak.

This highly plastic phenotype displays an unusual biogeographic distribution. It appears to range throughout Indonesia and the Philippines, but in the latter, it is completely absent in Cebu, where it is replaced by the preceding phenotype. An important question to ask is why is Cebu harboring a separate and unique phenotype? It’s unusual that a small, central region of the Philippines archipelago is capable of holding on to a potentially endemic “species” of Cirrhilabrus, although this phenomenon is repeatedly seen in other families, including certain species of gobies and dottybacks.



This variant is commonly encountered in the trade, with specimens collected from Bali and various other locations in Indonesia. This phenotype is reported to darken in captivity, usually losing its coloration pretty quickly. It also adopts the usual temminckii type temperament.
Cirrhilabrus cf. temminckii (Western Australia)

The Western Australian phenotype is essentially indistinguishable from the rest of the Indonesian population to the north. It, like the Indonesian and Philippine populations, possesses variably extensive black markings on the body and pectoral fin bases, which ranges from strongly present to completely absent.

Despite the phenotypic homogeneity between the Western Australian and Indonesian populations, it is possible that this phenotype may be harboring genetic drift due to the geographical isolation. This parallel can be seen in other Western Australian endemics, such as Cirrhilabrus randalli and Cirrhilabrus morissoni. The close proximity to Indonesia across the Timor Seas and the lack of any noticeable physical differences may suggest that this Western Australian population is a rather new arrival. Only time will tell if this phenotype will continue to speciate. As it is, the juveniles and females of the Western Australian phenotypes are slightly different, appearing to be mostly green instead of the usual terracotta.
This form is unobtainable in the trade due to lack of aquarium collection.

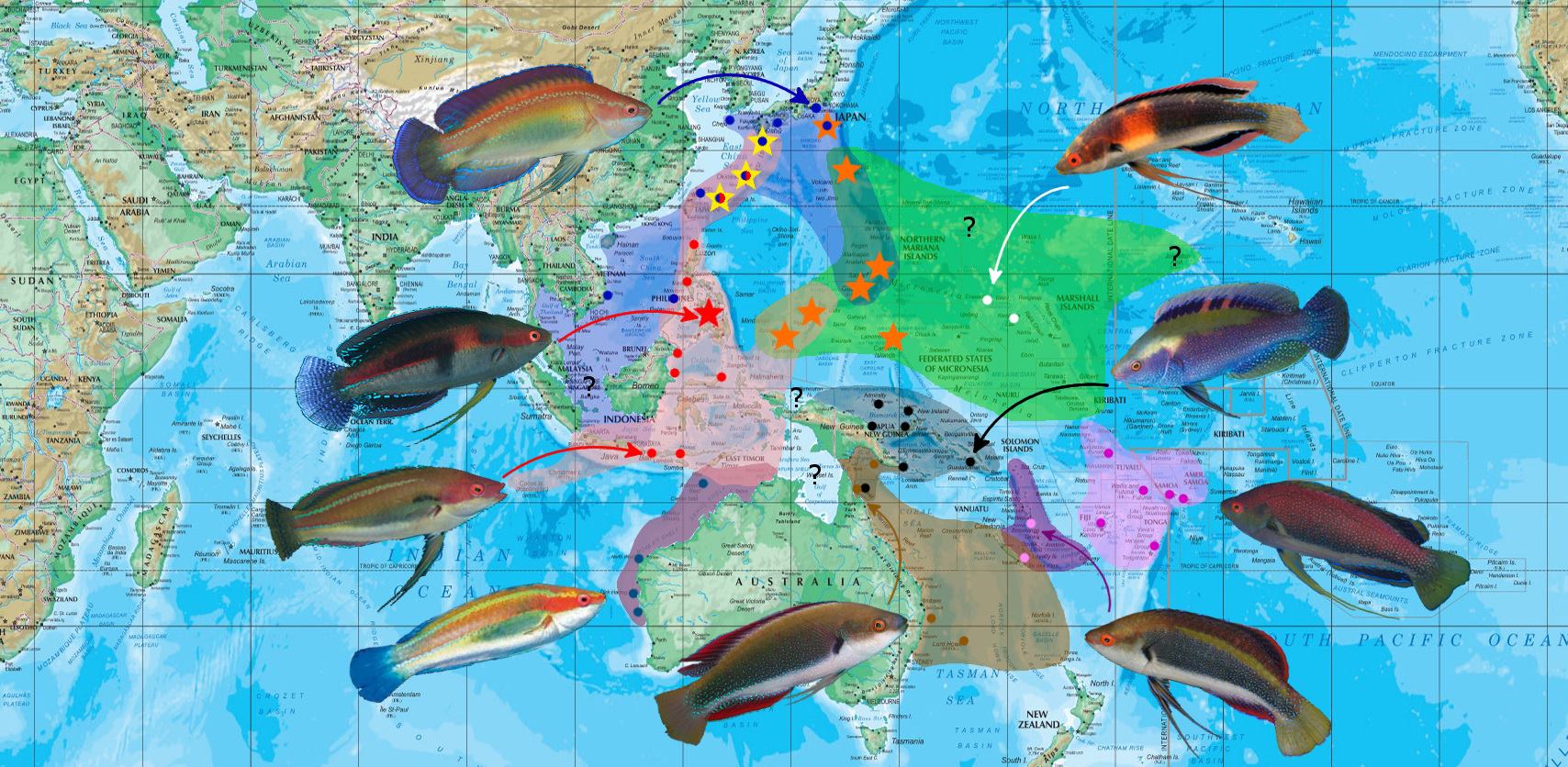
The map above summaries the approximate distribution of the temminckii species flock. Take note of the word approximate here. Due to the highly intractable nature of this group, caution has to be exercised in trying to make sense of this complex. Really, too little is known to make any concrete assumptions, and writing this article with our current limitations is not nearly enough to form a clear understanding of the group’s phylogenetics and biogeography.
In the later portion of this article, we’ll take a look at various anomalous examples, which muddles what little understanding we have of the group. Hybrids, aberrant distribution patterns and unknown phenotypes suggest the possibility of intra-species complexes within the currently known flock.
Again, take note of the Cebuan C. cf. temminckii and remember the features it shares with balteatus, in lacking the dorsal stripe and possessing the dorsal and anal fin medial stripes. Whether this phenotype is best associated with temminckii or balteatus is difficult to say within the limitations of this study.
The balteatus clade

Like the temminckii complex, the balteatus complex features members that are virtually inseparable via phenotype. However, unlike the former, the members here have been awarded full species level, highlighting yet again the inconsistencies in how the current classification treats distinct populations within Cirrhilabrus. Two currently accepted species comprise this flock — C. balteatus and C. katherinae. C. balteatus is restricted to the Marshall Islands, but C. katherinae presents us with a greater headache.
Cirrhilabrus balteatus

With its bright orange flank, this species is immediately recognizable from the temminckii complex. The ground coloration ranges from olivaceous, to gunmetal grey, to black, fading to a lighter shade, sometimes white, ventrally. The dorsum is thickly and rather prominently delimited in fir green, and the flank immediately behind the pectoral fin is branded in a highly prominent yellow-orange square.

C. balteatus possesses a pair of facial stripes tracing the upper and lower orbital limits. These, however, are often obfuscated and unobtrusive but can be traced along the edges of the face, which is brick red within the boundaries of these stripes. The dorsal and medial stripes are extremely vestigial and perforated, often times totally absent. The dorsal and anal fins are burgundy, and possess the characteristic medial horizontal stripes emarginated in blue.

The orange girdle of this species is very characteristic to the males. In nuptial display, this region lights up to a bright white. The typical facial and body stripes glow in an electrifying blue, but still remain somewhat nascent and obfuscated.
Cirrhilabrus balteatus is documented definitely from the Marshall Islands, but there are photos from Palau, which suggests the presence of this species, or at least a hybrid involving it. A record from the Caroline Islands has been documented, which is likely to be this species, but it is identified as C. katherinae. It is replaced by C. katherinae to the west. The two, like the temminckii group, presents a continuum in both biogeography and phenotype. They, however, have been weakly separated on the basis of phenotype.
C. balteatus is not common, but cannot be considered rare in the trade. The species is infrequently collected but does make an occasional appearance in fish stores. It is the most common Cirrhilabrus to be found in the Marshall Islands.
Cirrhilabrus katherinae

Cirrhilabrus katherinae is a polychromatic “species” with a bizarre geographical distribution. This, coupled with its phenotypic plasticity, may suggest that the “species” is actually of hybrid origin brought upon by neighboring species in their adjacent ranges. To begin, we’ll first have to look at the distribution ranges of this complex.
The map above shows the primary regions where Cirrhilabrus katherinae is found. Each region houses a distinct phenotype, but even within, spectrums of aberrant forms are documented, suggesting frequent hybridization and the inability of C. katherinae to maintain itself as a species.
Highlighted in red is the Ryukyu Arc, an island chain stretching from southern Kyushu to Taiwan. Osumi, Tokara, Amami, Okinawa, Miyako and Yaeyama bridge the gap in-between. Aparri, Cagayan, Northern Philippines, although not part of this arc group, is included in this portion of the range.
The Mariana Arc to the southeast is highlighted in purple. Guam and Saipan are the major landmarks that are taken into consideration here. Also included, although technically not part of the island arc grop, are Izu and Ogasawara to the north, and Yap to the south.
Finally, Palau is highlighted in green to the south, and here, another phenotype of C. katherinae is documented. These Island Arcs, including the Philippines archipelago, encircle the Philippine Sea.
It’s not possible to solve or even speculate the katherinae conundrum without extensive study. What we can do, however, is present the reader with various examples of phenotypic differences in the highlighted regions and hypothesize potential scenarios and parental lineages that may give rise to these forms.
Cirrhilabrus katherinae (Mariana Arc + Izu and Ogasawara)

The Mariana Arc phenotype for Cirrhilabrus katherinae is layered in three colors. The ground color for this form is orange-red, delimited strongly with a white ventral region. The dorsum is fir-green, running from the tip of the snout to the upper limit of the caudal peduncle. A black spot is present on the pectoral fin base. Like C. balteatus, the upper and lower orbital limits are traced with a pair of stripes, and these stripes extend to form the very vestigial dorsal and medial body stripes. The extrapolation of these stripes form the delimiting boundaries for the green, red and white body colorations.
The dorsal and anal fins are burgundy with a thick black medial stripe bordered in blue. The tail contains the usual scintilla, inscribed heavily in scrawling, and the pelvic fins are very long and trailing.




In full nuptial display, this phenotype lightens only on the posterior half. The barely noticeable dorsal and medial stripes in this portion of the body scintillate blue, and the ground color lightens to a downy white. Variable amounts of black develop on the white ventral region either as streaks or blotches.
The fins are clamped down, with only the anterior portion raised, and the caudal fin scintillates in a reflective blue. The nuptial coloration is another salient feature in distinguishing the various katherinae complex phenotypes.
The Mariana Arc is noted to receive fishes from Japan, Micronesia and rarely Hawaii, as strays. A number of species in this region are hypothesized to be of hybrid origin, such as Centropyge shepardi and Chaetodon flavocoronatus. Hybrid examples between C. ferrugata x C. loricula in the Philippines, and C. burgessi x C. declivis in the Marshall Islands have drawn similar parallels to the two aforementioned species in Guam, suggesting that this scenario may be in fact, entirely plausible. Dr. Richard Pyle seems to concur with this hypothesis, that “C. flavocoronatus may quite possibly be a hybrid derived species.”.
Could Cirrhilabrus katherinae in the Marianas share a similar background then? The Marianas Arc receives the occasional Cirrhilabrus temminckii and Cirrhilabrus balteatus from Japan and Micronesia respectively. Could these species be the precursor for C. katherinae? Again, we don’t know. Not without extensive study. Waifs of Japanese C. temminckii and/or Micronesian C. balteatus here are rare, therefore allowing the Marianas phenotype to achieve some form of reproductive isolation. Like many examples in the Mariana Arc, it is likely that this phenotype, given time, will eventually lead to complete speciation.
Another scenario could be that this was a more isolated and distinct “species” which has subsequently reconnected with one or both of its parents in recent times, and as such, is losing its distinctiveness.
Cirrhilabrus katherinae (Ryukyus Arc + Northern Philippines)

C. katherinae from the Ryukyu Arc displays the most variation, and occurs from Osumi in the Kagoshima prefecture, down to Aparri, Cagayan, at the northern tip of the Philippines. This phenotype is virtually inseparable from C. balteatus, and contains several additional features when compared against those of the Marianas Arc.
The ground coloration is essentially similar, being orange-red with cleanly delimited regions. The dorsum is entirely fir green from the snout to the upper tip of the caudal peduncle, and the ventral region is typically white. It however, possesses a light orange girdle in a fashion very similar to that of C. balteatus.




The placement and the extensiveness of the girdle contribute greatly to the variability of this “species”. Some specimens sport a thin girdle that is placed significantly distal to the pectoral fin base, almost midway through the body length. In others, the girdle is extensive enough to touch the pectoral fin base. Extreme examples may have the girdle entirely suffusing the anterior portion of the fish, forming a near bi-colored appearance. The posterior body is also very variable, ranging from reddish to entirely black.


This phenotype in nuptial display resembles C. balteatus extremely closely. The orange anterior saddle lightens to white, and the fish develops very weak blue scintilla on the dorsal and medial stripes. Unlike the Marianas phenotype, no lightening of the posterior is observed, both in the dorsal and medial stripes as well as the body.
Again, the highly variable phenotype might suggest a hybrid origin story. The Ryukyu Arc, however, presents a much greater complication than that of the Marianas. This region receives Cirrhilabrus temminckii, Cirrhilabrus cf. temminckii from the Philippines, and potentially, although rarely, Cirrhilabrus cf. temminckii from Cebu. If this is indeed a hybrid-originated species, determining the parental lineage is impossible without extensive, comprehensive and detailed genetic comparisons with every overlapping phenotype. To further complicate the situation, the Ryukyu Arc katherinae phenotype itself forms extensive hybrids with Cirrhilabrus temminckii in Japan, and other indecipherable mixes.

With so many sympatric phenotypes intermingling in the Ryukyus Arc, it’s unlikely that the katherinae form here can maintain reproductive isolation needed for legitimate speciation. It’s quite likely that this phenotype comprises of a cluster of hyper complex hybrids. We’ll take a look at some examples towards the end of this article.
Another interesting observation is the extreme similarity this phenotype has to Cirrhilabrus balteatus. While C. balteatus is presumably capable of reaching the Mariana Arc, it does not stray this far west to the Ryukyus, How or why this variant shows such similarity is another question to ponder about. Whether it has any genetic connection to C. balteatus is likewise unknown. If it does, could it be receiving balteatus DNA via katherinae from the Marianas or Palau?
Cirrhilabrus katherinae (Palau)

The katherinae phenotype from Palau resembles the preceding Ryukyus form rather closely, and is essentially inseparable. It, however, has a totally different nuptial pattern from those seen in the Mariana and Ryukyu Arcs. Its distribution in Palau means that the species cocktail found here is slightly different as well. Unlike the Ryukyus, this region may occasionally receive waifs of C. balteatus, as well as C. cf. temminckii from Cebu. It may also receive strays of the Marianas katherinae.


The nuptial coloration for this phenotype is dramatically different from the other two. In full nuptial display, the fins are clamped down except for the anterior third. The entire body turns white, save the dorso-anterior region just before the saddle.
Once again, assuming this form is indeed of hybrid origin, it simply is not possible to make out the parental lineage without extensive genetic testing. The Palauan and Ryukyu Arc phenotypes are biogeographically distinct from those in the Mariana Arc, and the “katherinae” in these two locations are unlikely to be anything more than hybrids, which strongly resemble that of the true katherinae. There are no other Mariana Arc endemics known outside of this range. Although we cannot conclusively confirm the parentage, we can be sure that the Palauan and Ryukyu Arc phenotypes are not katherinae.
The punctatus clade

The punctatus clade presents the reader with a more a classic and familiar case of allopatric speciation. Like the exquisitus complex, each biogeographic region of endemism houses a phenotypically distinct variant of the species, which could suggest incipient stages of speciation. At least one of these phenotypes has already been awarded full species level — Cirrhilabrus beauperryi.
Cirrhilabrus punctatus (Type – Coral Sea, Great Barrier Reef)

The type location for this species is One Tree Island, Great Barrier Reef, in Australia. This phenotype represents the type species for this complex, and all other forms of this species outside of this region should be addressed as Cirrhilabrus cf. punctatus due to varying differences in their appearance.

The species has a deep-olive green to brown dorsum, intensifying to a greenish grey or gunmetal medially before abruptly changing to white along the entire ventral region. Cirrhilabrus punctatus completely lacks any eye, dorsal or medial stripes, and is instead replaced by a constellation of fine metallic green or blue spots peppered extensively over its body.

The dorsal fins are red to burgundy, possessing the usual medial stripe, and likewise punctated in fine metallic spotting. The dorsal and anal fin stripes are a salient characteristic, which are less distinct and prominent than those in neighboring populations. The caudal fin is without any scrawling, is olive, and possesses the usual scintilla in a sidelight.

Despite the relative abundance of this species both in the trade and in the field, not many photos depicting this species in nuptial display exists. Judging from other members in this group, it is quite likely that some body whitening is to be expected during flashing.

The type representation for Cirrhilabrus punctatus ranges throughout the Coral Sea and the Great Barrier Reef. It strays very weakly into Papua New Guinea, where it overlaps with its allopatric sister, C. beauperryi. This unusual phenomenon works both ways, allowing C. beauperryi to cross into the Coral Sea region as well. This break in the allopatric boundary is thought to be fairly recent, and is explained later in the section discussing Cirrhilabrus beauperryi.
C. punctatus is common in the trade, appearing regularly from Australian exports. It is, however, fairly unpopular due to its large size, dull coloration and belligerent behavior.
Cirrhilabrus cf. punctatus (Vanuatu)

A different phenotype occurs eastwards in Vanuatu. This form is essentially identical to the population from the type locality and in the terminal males, an additional yellow posterior dorsal stripe tracing the lateral line. It delimits the olivaceous dorsum from the dusky medial region. Often times the dorsum may vary in color from the usual olive, to a more brick red or terracotta hue.

This phenotype is found only in Vanuatu. Vanuatu is an interesting location, in that although adjacent to Fiji in the eastern portion of Melanesia, it tends to harbor its own endemics not shared with the other island groups within this region. Cirrhilabrus bathyphilus and Cirrhilabrus exquisitus draw similar parallels, with each species being replaced by a distinct phenotype in Vanuatu and the other Melanesian Islands.
We are unable to find any photographs depicting this form in nuptial coloration, so aquarists are in a unique position in helping us fill this void. If you happen to have this phenotype in captivity, and have witnessed it in nuptial display, do contact us, preferably with a photo.
This phenotype is common in all Vanuatuan exports, and has the common name of “Port Vila Fairy Wrasse”, after Port Vila in Vanuatu. Like C. punctatus, it tends to gets aggressive in captivity and it has been reported to lose some of its coloration after time.
Cirrhilabrus cf. punctatus (Fiji)

The Fijian phenotype differs from the preceding two in lacking any dorsal stripe or color gradient in the dorsum. This form is slated rather extensively in a deep claret, obfuscating the underlying grey almost completely, but not without leaving a distinct margin to the dorsal coloration. The usual body spots are present, with an additional yellowish eye stripe running across from the upper lip to the edge of the gill plate. The dorsal and anal fin stripes, unlike those from the type locality, have the medial stripes distinct and outlined in blue.


In nuptial display, this phenotype lightens to a bright white over the entire dorsum, head and lower posterior ventral. The transverse eye stripe darkens from yellow to a deep olive. The dorsal fin is clamped down, except for the anterior half, which is held erect. The caudal fin displays varying levels of scintillation, and refracts in blues and green depending on the angle of incident light.
This phenotype is available regularly from Fijian imports, and like the other punctatus forms, is belligerent and bossy.
Cirrhilabrus beauperryi

Cirrhilabrus beauperryi is essentially just another punctatus variation. The only difference is that this phenotype has been split and awarded full species status. In C. beauperryi, the ground coloration is unusually colorful, ranging from blue to various shades of bright purple. The dorsum is copiously streaked in fir green, to olive, to ochre, and is messily delimited with jagged margins. The entire body, except for the dorsal patch, is heavily and intensely dotted in metallic blue or green.

The dorsal fin is hyaline, often tinted in yellow, and completely lacking in medial stripes. This is an important trait, because in every other geographical variant of punctatus, a longitudinal medial stripe is present. In C. beauperryi, however, a basal dorsal stripe is present, in dark purple, running throughout the dorsal fin. No other body or eye stripes are present.
All its median fins are heavily spotted in metallic blue, and its pelvic fins are exceptionally long and filamentous. It is worthy of mention that large, terminal males develop a strongly concave dorsal head profile.




This species shows little change in its nuptial display. Unlike C. punctatus, the dorsal fin in this species is held erect, while the caudal and anal fins are pressed against the body. The fins brighten to white, while the ochreous dorsum patch lightens in color. The body spots may also intensify in glitter.
Cirrhilabrus beauperryi ranges across Papua New Guinea and the Solomon Islands. Interestingly, this species is sympatric with its sister, Cirrhilabrus punctatus, in the Coral Sea and the Great Barrier Reef near Cairns. The two can be found together, although with C. beauperryi in much reduced numbers.
The vast difference in phenotypic appearance between C. punctatus and C. beauperryi suggests that the latter has gone the furthest in speciation. That is perhaps the reason why it was noticed by ichthyologists and subsequently split from C. punctatus, while the other more subtle phenotypes are still collectively regarded as one species.
Such a scenario makes for little sense in a sympatric overlap. The insurgence of C. beauperryi into the Australian region may be a relatively recent occurrence, with the forces once separating the two species removed, both fishes are now able to reconnect again. This leaky distribution works both ways, with C. punctatus also straying north into Papua New Guinea. Whether or not they hybridize is another question, but no hybrids have been documented so far and it’s likely that C. beauperryi has speciated far enough to maintain its genetic purity.
This species is uncommon in the aquarium trade, but is occasionally available from Australian imports.
Anomalies and hybrids

The complexity and intricacies of this group leaves us with a slew of unanswered questions in its wake, especially in the temminckii and katherinae complexes. We see multiple specimens spanning an entire spectrum of variability between each of the known phenotypes, suggesting that frequent hybridization is taking place. The hybrids also go to show that we don’t quite fully understand the distribution and biogeography of this species complex yet, with some specimens expressing phenotypic qualities of various “species” that are not documented in that particular range.


An example of a hybrid scenario is pictured above in Japan. Compare the type Cirrhilabrus temminckii in the background to the fish in the foreground. The complete and prominent longitudinal medial stripe in the dorsal and anal fin suggests that this fish has Ryukyus katherinae DNA. However, the complete and prominent dorsal stripe suggests that temminckii might also be involved. Whether this is an F1 hybrid between the two, or a result of a more complicated mix is something we can’t concretely answer. Remember, the Ryukyu Arc is a literal soup of temminckii group members, and with so many species and hybrids mixed together, pinpointing the parental lineage without genetic testing is impossible.

Yet another anomalous example is seen here. The cleanly delimited coloration recalls that of the Mariana Arc examples of katherinae. However, this specimen was collected from Aparri, northern Philippines. The northern Philippines is home to the Ryukyus katherinae phenotype, so whether or not this is a hybrid involving that remains a mystery.

Another unexplainable phenotype occurs in Cebu. As far as we know, this region is presumably entirely replaced by the Cebuan Cirrhilabrus cf. temminckii with the rotated “L” medial stripe. The darkened posterior above recalls that of the Ryukyus katherinae, especially those coming from Aparri in the northern Philippines. However, fish from that geographical region have never been recorded this far south in Cebu, reaching Luzon as their southernmost limit.
If this hybrid indeed contains katherinae DNA, could some have made it to Cebu? If not, then what could it possible be? The Cebuan temmcinkii variant resembles balteatus and katherinae more closely with regards to its, but again, too many uncertainties and answered questions.

The temminckii group is perhaps single handedly the most complex and intractable of the Cirrhilabrus groups. Going further than speculative hypothetic relationships is a stretch, and extensive study needs to be done to answer some of these questions. Even with thorough genetic analysis, it is unlikely that any real differences can be compared at this stage. The temminckii and katherinae clade members are too vastly amalgamated, which may stem from their incipient evolutionary divergence coupled with their wide sympatric overlay. It would be interesting to revisit this group in a couple hundred thousand, maybe a million years later to check on their divergent progress. Perhaps the group will eventually homogenize. We simply don’t know.
The punctatus clade is comparatively more straightforward, and there we see a typical phenotypic diversity attributed to allopatric separation.
In summary, this article should not be seen as an answer to the temminckii group, but rather as a guide to the questions that need answering. Only three groups are left before the series comes to an end — rubriventralis, filamentosus and laboutei.
