
At last, we’ve reached the thrilling conclusion to this comprehensive review of Fairy Wrasse evolution, and it’s time to once more dwell upon the topic of the group’s origins. As we’ve previously discussed, this enormously diverse genus can be comfortably categorized into a pair of major lineages that serve to organize these fishes into more comprehensible portions.
There are species with short pelvic fins, and there are species with longer pelvic fins. Corresponding with this difference are numerous other similarities regarding their respective color patterns, particularly when it comes to the reflective caudal fin of the long-finned group. Female coloration can likewise be informative, as this tends to be a conserved feature within and across the various species groups. But what all of these species have shared up until now is a fairly standard morphology.

By and large, the morphological counts of things like fin spines and rays, lateral line scales and gill rakers are homogenous across the entire genus. Certain individual groups are recognizable due to some minor alteration of their proportions (e.g. the pointy head seen in the lubbocki or filamentosus groups), but the overall morphology is still much the same. There is, however, still one more species remaining which we have yet to cover, and it differs from its relatives in somewhat more meaningful ways.

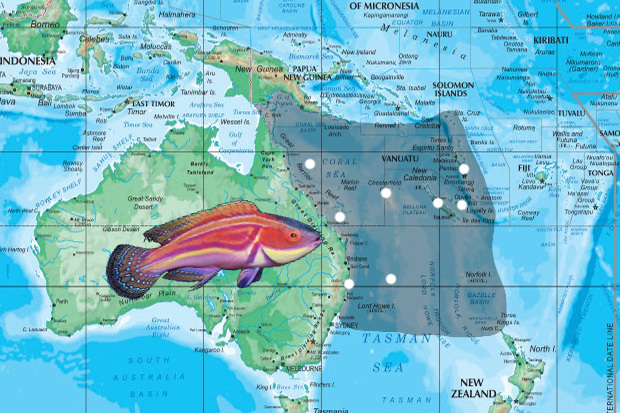
Cirrhilabrus laboutei is a fairy wrasse sui generis. It was first made known in a 1976 guide to the fishes of New Caledonia authored in part by Pierre Laboute, who included a photograph and a short sentence mentioning its size (15cm… though it’s actually closer to 12cm), habitat depth (15-30m), and the observation that it hovers several meters above the bottom along with an unidentified fairy basslet.
Mr. Laboute, who went on to author a number of guides to the New Caledonian fauna, shared information of his find with Dr. John Randall, which led to its official scientific description in 1982. Pierre, for his efforts, is now immortalized with one of the more resplendent wrasses in the sea—Laboute’s Fairy Wrasse.

Despite still having essentially the same morphological counts as other fairy wrasses, two features unique to this fish serve to set it apart from its congeners. The first has to do with the unusually elongated spines of the anal fin, something seemingly not found anywhere else in the entire family. These only develop in large males, and they are erected and spread apart during the nuptial display. Whether these finger-like spines serve any tactile function or are merely visual stimulants for the females is entirely unknown.
Equally idiosyncratic are the rounded pelvic fins, which again have no real parallel within this genus; however, the closely related flasher wrasses (Paracheilinus) share this fin shape. Other related groups—Pseudocheilinus, Pseudocheilinops, Pteragogus—possess similarly rounded pelvic fins, lending support to the notion that the elongate fins seen in most fairy wrasses are a derived feature. This can be taken as further evidence that C. laboutei likely belongs to an early lineage which predates the acquisition of this trait.

Other characteristics also stand out as unusual. Take, for instance, the great similarity between the coloration of juveniles and females relative to mature males (other species typically show a fairly drastic change during development). We do still see some of the same juvenile landmarks found elsewhere in the genus, namely the three prominent lines running along the length of the body.
This is not so different from the juveniles of the lunatus, jordani or rubriventralis groups, but laboutei is noteworthy in that these juvenile lines don’t ever fade away with age as we would expect. The upper two are kept into adulthood and serve to border a stripe extending posteriorly from the eye, while the lowermost line remains associated with the posterior lateral line scales.

If we compare the arrangement of laboutei’s patterning to its relatives, we see a mix of features from a variety of different lineages. The aforementioned eye stripe is a common trait in the diverse jordani group, while the short dashes present near the pectoral fin base appear again in the distantly related C. exquisitus and C. johnsoni. The caudal fin, delicately patterned in a series of concentric lines and dots, is not so different from the temminckii or rubriventralis groups, but, importantly, laboutei’s fin never has any indication of a reflective sheen to it.

Mature males develop a characteristic black band dorsally along the head, which we can only find vague similarities to in things like lubbocki, jordani and rubrimarginatus. Again, the narrative here is that of a feature which is common to many distantly related groups and which laboutei has put its own idiosyncratic twist on.
The nuptial pattern is poorly documented and in many ways unknown. Certain specimens show a blackening of the dorsal and anal fins and an intensification of the colors along the body, but these may merely represent individual variation within the species. Size and biogeography don’t appear to correlate with these differences in male coloration, as similarly sized males from both Australia and Vanuatu show a range of phenotypic variations.
The most plausible inference that can be made is that the kind of shimmer in the stripes or caudal fin that is otherwise so prevalent in fairy wrasses is mostly or entirely lacking in laboutei. Might this indicate yet another ancestral trait? Aquarists could serve an important role in advancing our understanding of this species by recording the displays of captive specimens!
Since its initial discovery, C. laboutei has turned up in other nearby regions of the Southwest Pacific, including Vanuatu, the Loyalty Islands, the Chesterfield Islands, Elisabeth Reef, and as far north as Cairns along the Great Barrier Reef and as far south as the Solitary Islands in New South Wales. This is a virtually identical biogeography to C. bathyphilus, but, unlike with that fish, there doesn’t appear to be a sister species found to the east. Poor C. laboutei is all alone in this world—the only Cirrhilabrus in such phylogenetic isolation.

Aquarium specimens are liable to come from any number of locales: Vanuatu, New Caledonia, Australia. Specimens collected near Cairns, Australia are said to range from 10-40 meters, with most found in the 10-20 meter range. The fish collectors and exporters in this region do a far better job than what we generally see from their Indonesian and Philippines counterparts, and so specimens of laboutei typically arrive at their destination in tip-top shape. For such a robustly sized Cirrhilabrus, this species is rather on the wimpy side and is far more likely to be victimized by boisterous tankmates than to cause any problems itself. Pierre Laboute’s wrasse is a lover, not a fighter, though whether Pierre Laboute himself was of a similar disposition I cannot speak to.

When we began this journey into bettering our knowledge of Cirrhilabrus evolution, we presented a pair of competing hypotheses regarding the basalmost member of the genus—what one might colloquially refer to as the most “primitive” living species. “Primitive” is a bit of a loaded term here, a pejorative which unnecessarily implies a less-evolved state of affairs. In the case of Cirrhilabrus, it has become increasingly obvious through the course of this study that C. laboutei has that special blend of shared, ancestral characteristics and unique features which mark it as different from the rest. But even though, as we suggest, it occupies its own isolated lineage with respect to its congeners, remember that all fairy wrasses share the same common ancestor at some point in their past. Cirrhilabrus laboutei may very well belong to a relictual lineage from early in the evolution of the genus, but, in a sense, so do all the other species. It’s the relative diversification and speciation which marks these two basalmost clades as different. Perhaps a few tens of millions of years ago it would have been the laboutei clade which dominated the oceans?

As a final point concerning fairy wrasse evolution, we present here the most comprehensive molecular study done to date on the genus. This isn’t saying much, though, as there has yet to be any real effort by the scientific community to review Cirrhilabrus using the many molecular tools at our disposal. How this state of affairs has been allowed to go on for so long is beyond me, but hopefully this nascent attempt will spur further inquiry. This tree is based on publicly available mitochondrial CO1 data, and, for the most part, it conforms to what we had previously arrived at from morphological and biogeographical study alone. There are minor discrepancies with the arrangement of some of the long-finned groups in the CO1 tree, but there is too little data to draw any meaningful conclusions. What is most urgently needed is the inclusion of C. laboutei into this dataset, to hopefully corroborate its placement basally in the genus.
And with that, we have finally reached the end of this voyage into the uncharted waters of Cirrhilabrus, perhaps with just as many questions now as when we first started. What should be clear is just how imperfect and incomplete our knowledge truly is—how many taxonomic inconsistencies plague this genus. There are many species yet to be named, and there may yet be more awaiting discovery in the briny depths of some obscure corner of the ocean. It will undoubtedly be a great long while before enough morphological and molecular research is published to confirm or refute what we’ve put forth in this long-winded review, and we certainly do not envy whomever attempts to at last elucidate the true diversity hiding within taxonomic nightmares like C. temminckii and exquisitus.
Addendum
Several articles have been updated with new information:
New photo of Cirrhilabrus lanceolatus was added.
New photos of Cirrhilabrus claire were added.
New photo of Cirrhilabrus bathyphilus was added.
Full nuptial coloration of Cirrhilabrus bathyphilus documented and updated.
Terminal male phase of Cirrhilabrus cf. solorensis documented and updated.
New photos of Cirrhilabrus cyanopleura x C. cf. solorensis hybrids were added.
New photos of Cirrhilabrus exquisitus from Africa were added.
New photo of C. cf. exquisitus from Sumatra was added.
Cirrhilabrus rubrimarginatus updated with a range extension into New Caledonia.
New video displaying caudal scintilla in Cirrhilabrus katoi was added.
Acknowledgments

A tip of the hat to my indefatigable co-author, Lemon, who took it upon himself to pen most of this series. Several others have been instrumental along the way: Hiroyuki Tanaka, Gerry Allen, Rudie Kuiter, Kevin Kohen, Eli Fleishauer, Vincent Chalais, Brian Greene and the Japanese divers who have documented these fishes in situ.
